International Journal of Systematic and Evolutionary Microbiology (2008), 58, 919–923
Bacillus coahuilensis sp. nov., a moderatelyhalophilic species from a desiccation lagoon in theCuatro Cie´negas Valley in Coahuila, Mexico
Rene´ Cerritos,1 Pablo Vinuesa,2 Luis E. Eguiarte,1 Luis Herrera-Estrella,3Luis D. Alcaraz-Peraza,4 Jackeline L. Arvizu-Go´mez,4 Gabriela Olmedo,4Enrique Ramirez,4 Janet L. Siefert5 and Valeria Souza1
1Departamento de Ecologı´a Evolutiva, Instituto de Ecologı´a, Universidad Nacional Auto´noma de
Me´xico, Apartado Postal 70-275, Me´xico D.F. 04510, Mexico
2Programa de Ingenierı´a Geno´mica, Centro de Ciencias Geno´micas, Universidad Nacional
Auto´noma de Me´xico, Apartado postal 565-A, Cuernavaca, Mor. 62210, Mexico
3Langebio, Cinvestav, Apartado Postal 629, Irapuato, Gto. 36821, Mexico
4Departamento de Ingenierı´a Gene´tica de Plantas, Cinvestav Unidad Irapuato, Apartado Postal 629,
5Department of Statistics, Rice University, Houston, TX 77251, USA
A moderately halophilic, Gram-positive and rod-shaped bacterium, strain m4-4T, was isolatedfrom a Chihuahuan desert lagoon in Cuatro Cie´negas, Coahuila, Mexico. Strain m4-4T was foundto grow optimally at 30–37 6C, pH 7.0–8.0 and 5 % NaCl and to tolerate from 0.5 % to 10 %NaCl. It was shown to be aerobic. The genomic DNA G+C content was about 37 mol%. Strainm4-4T exhibited minimal or no growth on most sugars tested. Its major cellular fatty acids wereC14 : 0, C16 : 0 and C18 : 1. Based on phylogenetic analysis of 16S rRNA and recA genesequences, we observed that the closest relatives of the isolate are moderately halophilic Bacillusspecies, with 16S rRNA gene sequence similarity ranging from 96.6 to 97.4 % (Bacillusmarisflavi, Bacillus aquimaris and Bacillus vietnamensis). Additionally, using genomic data it wasdetermined that the type strain contains a total of nine rRNA operons with three slightly differentsequences. On the basis of phenotypic and molecular properties, strain m4-4T represents a novelspecies within the genus Bacillus, for which the name Bacillus coahuilensis sp. nov. isproposed, with the type strain m4-4T (5NRRL B-41737T 5CECT 7197T).
A number of halophilic and moderately halotolerant,
2006). In this study, the Bacillus strain m4-4T was isolated
Gram-positive, endospore-forming aquatic isolates in the
in August 2003 from a desiccation lagoon in the Churince
genus Bacillus have been described. A large number of
system, a hydrological system on the western side of the
them have been isolated from marine environments
Cuatro Cie´negas Valley in Coahuila, Mexico (26u 50.8309N,
(Siefert et al., 2000; Yoon et al., 2003, 2004; Noguchi
et al., 2004; Yoon & Oh, 2005; Lee et al., 2006). However,
Strain m4-4T was analysed using taxonomic and biochem-
little is known about species inhabiting non-marine, high
ical methods. Two markers were used for phylogenetic
salinity aquatic environments (Lim et al., 2006; Souza et al.,
reconstruction (16S rRNA and recA gene sequences). Studies have shown that more robust results are obtained
GenBank/EMBL/DDBJ accession numbers for the 16S rRNA gene
when additional markers such as housekeeping genes are
sequences: m4-4T, EF014450, EF014451 and EF014452; and the
used, especially in closely related isolates (Stackebrandt
recA gene sequences: m4-4T, EF014455; B. marisflavi TF-11,
et al., 2002; Zeigler, 2003). We determined the phylogenetic
EF014457; B. vietnamensis NRIC 0530, EF014458; B. vietnamensis
affiliation of the isolate m4-4T by means of 16S rRNA gene
NRIC 0531T, EF014459; B. vietnamensis NRIC 0532, EF014460; B.
phylogeny reconstruction and determined its taxonomic
status as a representative of a novel species by using a
Photomicrographs of Bacillus coahuilensis m4-4T, a phylogenetic tree of
polyphasic approach. The study also included genomic
the recA sequences of m4-4T and other bacilli and a table showing thefatty acid composition of m4-4T are available with the online version of
analysis to determine environmental genome size and
Strain m4-4T was isolated from surface water samples that
1500 uC for 3 min, increasing at the rate of 40 uC min21 to
were taken and placed in sterile flasks. These were subjected
a final temperature of 3000 uC, which was maintained for
to a shock temperature of 80 uC for 20 min by means of
20 min. Helium was used as carrier gas with a constant
damp heat (Istock et al., 2001). Subsequently, 1 : 100 and
flow of 1 ml min21. Fatty acid methyl esters were identified
1 : 1000 dilutions were made. Aliquots (100 ml) from each
using the mass spectral library search (NIST MS Data Base)
dilution, as well as from the original water samples, were
distributed by the National Institute of Standards and
placed in Petri dishes with marine agar 2216 medium (MA;
Difco) and incubated at 37 uC for 2 days. Cultures were
A combination of Sanger (Nunally, 2005) and 454 Life
purified by subculturing on the same medium and
Sciences sequencing methods (Margulies et al., 2005) were
maintained at –80 uC in 5 % MA and 15 % (w/v) glycerol.
used to sequence the m4-4T genome, as described by
We studied the cell morphology and sporulation process
Alcaraz et al. (2008). The genome sequencing found nine
for strain m4-4T using phase-contrast microscopy. Cells
ribosomal operons; three of them had slight differences,
were negatively stained with 1 % (w/v) malachite green and
giving sequences m4-4a, b and c (Fig. 1). The G+C
contrasted with 1 % (w/v) safranine. Characterization of
content was obtained directly by genomic analysis.
strain m4-4T included the study of cultural, physiological
The 16S rRNA gene was amplified using the 27F and 1492R
and biochemical parameters. Single carbon source assim-
primers under conditions described previously (Lane,
ilation tests were performed in MA (4 g l21), replacing the
1991) in 100 ml final volume. The recA gene was chosen
yeast extract and peptone with the main carbon source.
for sequencing and phylogenetic analysis. Oligonucleotide
Nitrate reduction was determined as described by La´nyı´
primers were designed using the recA genes of the complete
(1987) in the presence and absence of 3 % (w/v) NaCl.
genomes of Bacillus strains reported in GenBank. These
Growth at different temperatures was measured on MA
primers extended from position 28 to 48 (59-GATCG-
between 30 and 50 uC. Urease activity was determined as
TCARGCAGSCYTWGAT-39) and from position 583 to
described previously by Cowan & Steel (1965).
602 (59-TTWCCRACCATAACSCCRAC-39), yielding a
For quantitative analysis of whole-cell fatty acids, strain
574 bp product. PCR mixtures (25 ml) were prepared with
m4-4T was cultivated on MA for 2 days at 37 uC. The
1 U Taq polymerase (Roche), 2.5 mM MgCl2, 1 mM
whole-cell fatty acid composition was determined by using
dNTPs, 2 mM each recA primer and 1 ml DNA (25–
a gas chromatograph (model 5890; Hewlett Packard)
100 ng ml21). The PCR program was one cycle of initial
equipped with a capillary column HP-5MS (30 m6
denaturation at 95 uC for 5 min, 30 cycles of denaturation
0.25 mm i.d.; 0.25 mm film thickness) coupled to a mass
at 95 uC for 30 s, annealing at 45 uC for 30 s and extension
spectrometer detector (model 5972; Hewlett Packard).
at 72 uC for 60 s, and a final extension cycle at 72 uC for
Operating conditions were an injection temperature of
5 min. PCR products were purified using a gel extraction
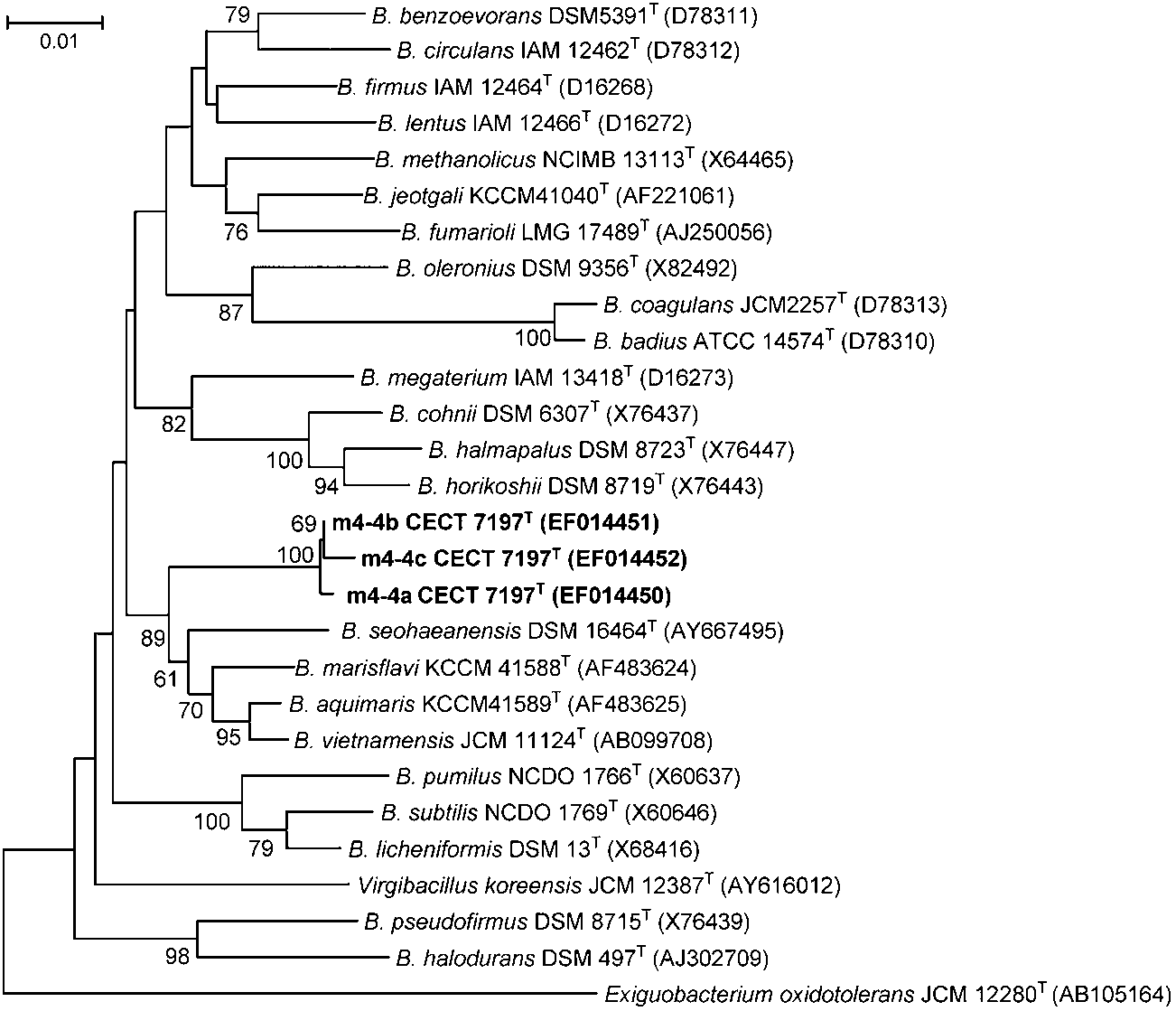
Fig. 1. Unrooted phylogenetic tree using theneighbour-joining method and derived from theanalysis of the 16S rRNA gene sequences ofstrain m4-4T and other representative Bacillusstrains. Numbers next to the branches repres-ent bootstrap values expressed as percentagesof 2500 replications; only values greater than70 % are indicated. GenBank accession num-bers of sequences are shown in parentheses. Bar, 0.01 substitutions per nucleotide position.
International Journal of Systematic and Evolutionary Microbiology 58
DNA kit (Qiagen). For the 16S rRNA gene, a fragment of
sequences obtained with the Ribosomal Database Project
approximately 1400 bp was sequenced with the primer set
and the NCBI databases. Phylogenetic reconstruction for
reported previously (Sacchi et al., 2002). For the recA gene,
the recA gene was done using seven complete genomes of
a 450 bp segment was sequenced from strain m4-4T, B.
Bacillus strains reported in the NCBI database. Sequences
marisflavi TF-11T and from four isolates of B. vietnamensis
were aligned using the CLUSTAL_W program (Thompson
(NRIC 0530, 0531T, 0532 and 0533). The sequencing
et al., 1994). Phylogenetic reconstruction for the 16S rRNA
reaction had a total volume of 15 ml consisting of 2 ml Big
and recA genes was done using the neighbour-joining
Dye Terminator sequencing buffer (Applied Biosystems),
algorithm with Kimura two-parameter distances, as
1.6 mM primer and 5 ml purified amplified product. The
implemented in MEGA3 (Kumar et al., 2004).
amplification conditions were as follows: one cycle of5 min at 95 uC, and 45 cycles of 10 s at 95 uC, 10 s at 50 uC
Strain m4-4T was subjected to morphological and physio-
and 4 min at 60 uC. Sequencing was done in a capillary
logical tests that showed significant differences with respect
sequencer (ABI-Avant 100). Sequences (GenBank accession
to other closely related Bacillus species (Table 1). Strain
numbers EF014450–EF014452, EF014455 and EF014457–
m4-4T grew on only three carbon sources (starch, glycerol
EF014461) were manually edited with the BioEdit program
and trehalose). Cells were rod-shaped, approximately 0.5–
(Hall, 1999). In the case of the 16S rRNA gene sequences,
0.7 mm in diameter and 1.5–3 mm in length after 2 days of
isolate identities were established by comparing the
cultivation at 37 uC (Supplementary Fig. S1, available in
Table 1. Differential characteristics of strain m4-4T and closely related strains
Strains: 1, m4-4T; 2, B. marisflavi JCM 11544T; 3, B. aquimaris JCM 11545T; 4, B. vietnamensis NRIC 0531T. The four strains were positive forutilization of starch, glycerol, L-glutamine, citrate, trehalose and fumarate. All strains were negative for nitrate reduction, H2S and urease.
*C, Central; S, subterminal. DLY, Light yellow; O, orange; PO, pale orange; PY, pale yellow. di, iso; ai, anteiso.
IJSEM Online). The G+C content of 37 mol% for strain
Description of Bacillus coahuilensis sp. nov.
m4-4T is significantly different from that for B. marisflavi
Bacillus coahuilensis (co.a.hui.len9sis. N.L. masc. adj.
(49 mol%) and B. vietnamensis (43–44 mol%), but not
coahuilensis in reference to Coahuila, the state in Mexico
from that for B. aquimaris (38 mol%).
where the type strain was collected).
The major cellular fatty acids of strain m4-4T were C14 : 0
Vegetative cells are rod-shaped, occurring in large chains
(29.4 %), C16 : 0 (22.3 %), C18 : 1 (15.2 %) and C17 : 0 (7.9 %).
(Supplementary Fig. S1a), approximately 0.5–0.7 mm in
Fatty acids occurring in minor amounts were C12 : 0
diameter by 1.5–3 mm in length. Central ellipsoidal
(1.3 %), anteiso-C17 : 0 (4.7 %) and anteiso-C15 : 0 (4.8 %)
endospores are observed in swollen sporangia and are
(Supplementary Table S1). Fatty acids profile comparisons
1.0 mm wide and 1.5–1.7 mm long (Supplementary Fig. S1b,
between strain m4-4T and other species of the genus
c). Colonies on MA are light yellow and 2–5 mm in
Bacillus reveal significant differences (Table 1).
diameter after 2 days growth at 37 uC; they are low, convex,
16S rRNA gene sequence similarity between strain m4-4T
circular and slightly irregular. Optimal growth temperature
and type strains of other phylogenetically closely related
is 30–37 uC and the maximum growth temperature is 45 uC.
Bacillus species (B. marisflavi, B. aquimaris and B.
Minimum pH for growth lies between 5.0 and 5.5, the
vietnamensis) ranged from 96.6 to 97.4 %. Values obtained
optimum pH for growth is between 7 and 8 and the
in this study meet widely accepted criteria for delineating
maximum pH for growth is 9. Acid is produced from
species in current bacteriology (Stackebrandt & Goebel,
glycerol, but not from D-glucose or lactose. Citrate and
1994). A 16S rRNA gene-sequence-based neighbour-join-
fumarate can be utilized. Nitrate reduction was not present.
ing phylogeny analysis revealed that the three different
H2S and urease are not produced. Does not utilize sucrose,
ribosomal operons of strain m4-4T formed a tight and
lactose, arabinose, dulcitol, fructose, adonitol, D-sorbitol,
highly supported clade (100 % bootstrap support) nested
salicin, D-mannitol, D-xylose, L-rhamnose and L-glutamine
within a deeper cluster that comprises B. aquimaris, B.
as sole carbon and energy sources. DNA G+C content of
marisflavi, B. vietnamensis and Bacillus seohaeanensis at a
the type strain is 37 mol%. Halotolerant, growing in NaCl
bootstrap confidence level of 87 % (Fig. 1). In addition, a
salt concentration from 0.5 to 10 %. The major fatty acids
recA-based neighbour-joining tree also grouped strain
are C14 : 0, C16 : 0 and C18 : 1. Additionally, based on genome
m4-4T as a strongly supported monophyletic lineage
analysis, strain m4-4T showed nine ribosomal operons with
(Supplementary Fig. S2), which is distinct from the clade
comprising B. marisflavi and B. vietnamensis.
The type strain, m4-4T (5NRRL B-41737T 5CECT 7197T),
Our results show that strain m4-4T can grow in medium
was isolated from a desiccation lagoon in the Cuatro
containing NaCl in the range 0.5 to 10 % (w/v). From these
Cie´negas Valley in Coahuila, Mexico.
data we concluded that this Bacillus strain is moderatelyhalophilic (Ventosa et al., 1998).
In this study we described a Bacillus isolate using
This research was supported by a CONACyT scholarship to C. R. The
biochemical and genomic data as well as phylogenetic
project was funded by SEMARNAT/CONACyT and SEP CONACyT
reconstructions involving 16S rRNA and recA gene
(C01-0237/A1 and 44673 Q) to V. S. and L. E. F. We thank Antonio
sequences. This approach showed that m4-4T is a member
Cruz, Laura Espinosa and Jose´ Luis Herna´ndez for specialized
of a distinct group within the genus Bacillus. The strain
technical assistance and Morena Avitia and Miguel Contreras for
displayed characteristics typical of Bacillus species, like
laboratory work. Special thanks to Alejandro Rooney for incorporat-
spore production and low DNA G+C content (37 mol%).
ing the strains to the NRRL collection and Luisa Falcon and Ana M. Noguez for thoughtful comments and Mark Schneegurt for providing
However, the fatty acid composition for strain m4-4T is
completely different from those of other closely relatedBacillus species (Supplementary Table S1). Chains C14, C16,and
C18 are characteristic for this novel isolate.
Phylogenetic analysis using 16S rRNA gene sequencesshowed that the novel isolate formed a distinct clade
Alcaraz, L. D., Olmedo, G., Bonilla, G., Cerritos, R., Herna´ndez, G.,
compared with the closely related type strains of B.
Cruz, A., Ramı´rez, E., Putonti, C., Jime´nez, B. & other authors (2008). The genome of Bacillus coahuilensis reveals adaptations essential for
marisflavi (JCM 11544T), B. aquimaris (JCM 11545T), B.
survival in the relic of an ancient marine environment. Proc Natl Acad
vietnamensis (NRIC 0531T) and B. seohaeanensis (DSM
16464T). Phylogenetic reconstruction using recA gene
Cowan, S. T. & Steel, K. J. (1965). Manual for the Identification of
sequences also showed that the novel isolate formed a
Medical Bacteria. Cambridge: Cambridge University Press.
distinct group compared with B. marisflavi JCM 11544T
Hall, T. A. (1999). BioEdit: a user-friendly biological sequence
and B. vietnamensis strains NRIC 0531T, 0530, 0532, and
alignment editor and analysis program for Windows 95/98/NT.
0533. We suggest, on the basis of the data described above,
that strain m4-4T should be placed within the genus
Istock, C. A., Ferguson, N., Istock, N. L. & Duncan, K. E. (2001).
Bacillus as a representative of a novel species, for which the
Geographical diversity of genomic lineages in Bacillus subtilis
name Bacillus coahuilensis sp. nov. is proposed.
(Ehrenberg) Cohn sensu lato. Org Divers Evol 1, 179–191.
International Journal of Systematic and Evolutionary Microbiology 58
Kumar, S., Tamura, K. & Nei, M. (2004). MEGA3: integrated software
Souza, V., Espinosa-Asuar, L., Escalante, A. E., Eguiarte, L., Farmer, J.,
for molecular evolutionary genetics analysis and sequence alignment.
Forney, L., Lloret, L., Rodriguez-Martı´nez, J. M., Soberon, X. & other
authors (2006). An endangered oasis of aquatic microbial diversity in
Lane, D. J. (1991). 16S/23S rRNA sequencing. In Nucleic Acid
the Chihuahuan desert. Proc Natl Acad Sci U S A 103, 6565–6570.
Techniques in Bacterial Systematics, pp. 115–175. Edited by E.
Stackebrandt, E. & Goebel, B. M. (1994). Taxonomic note: a place
Stackebrandt & M. Goodfellow. Chichester: Wiley.
for DNA-DNA reassociation and 16S rRNA sequence analysis in
La´nyı´, B. (1987). Classical and rapid identification methods for
the present species definition in bacteriology. Int J Syst Bacteriol 44,
medically important bacteria. Methods Microbiol 19, 1–67.
Lee, J.-C., Lim, J.-M., Park, D.-J., Jeon, C. O., Li, W.-J. & Kim, C.-J.
Stackebrandt, E., Frederiksen, W., Garrity, G. M., Grimont, P. A. D.,
(2006). Bacillus seohaeanensis sp. nov., a halotolerant bacterium that
Ka¨mpfer, P., Maiden, C. J. M., Nesme, X., Rossello´-Mora, R., Swings, J.
contains L-lysine in its cell wall. Int J Syst Evol Microbiol 56,
& other authors (2002). Report of the ad hoc committee for the re-
evaluation of the species definition in bacteriology. Int J Syst EvolMicrobiol 52, 1043–1047.
Lim, J. M., Jeon, C. O., Lee, S. M., Lee, J. C., Xu, L. H., Jiang, C. L. &Kim, C. J. (2006). Bacillus salarius sp. nov., a halophilic, spore-
Thompson, J. D., Higgins, D. G. & Gibson, T. J. (1994). CLUSTAL W:
forming bacterium isolated from a salt lake in China. Int J Syst Evol
improving the sensitivity of progressive multiple sequence alignment
through sequence weighting, position-specific gap penalties andweight matrix choice. Nucleic Acids Res 22, 4673–4680.
Margulies, M., Elgholm, M., Altman, W. E., Attiya, S., Bader, J. S.,Bemben, L. A., Berka, J., Braverman, M. S., Chen, Y. J. & other
Ventosa, A., Nieto, J. & Oren, A. (1998). Biology of moderately
authors (2005). Genome sequencing in microfabricated high-density
halophilic aerobic bacteria. Microbiol Mol Biol Rev 62, 504–544.
picolitre reactors. Nature 437, 376–380.
Yoon, J.-H. & Oh, T.-K. (2005). Bacillus litoralis sp. nov., isolated from
Noguchi, H., Uchino, M., Shida, O., Takano, K., Nakamura, L. K. &
a tidal flat of the Yellow Sea in Korea. Int J Syst Evol Microbiol 55,
Komagata, K. (2004). Bacillus vietnamensis sp. nov., a moderately
halotolerant, aerobic, endospore-forming bacterium isolated from
Yoon, J. H., Kim, I. G., Kang, K. H., Oh, T. K. & Park, Y. H. (2003).
Vietnamese fish sauce. Int J Syst Evol Microbiol 54, 2117–2120.
Bacillus marisflavi sp. nov. and Bacillus aquimaris sp. nov., isolated
Nunally, B. K. (2005). Analytical Techniques in DNA Sequencing. Boca
from sea water of a tidal flat of the Yellow Sea in Korea. Int J Syst Evol
Raton, FL: Taylor & Francis CRC Press.
Sacchi, C. T., Whitney, A. M., Mayer, L. W., Morey, R., Steigerwalt, A.,
Yoon, J.-H., Kim, I.-J., Kang, K. H., Oh, T.-K. & Park, Y.-H. (2004).
Boras, A., Weyant, R. S. & Popovic, T. (2002). Sequencing of 16S
Bacillus hwajinpoensis sp. nov. and an unnamed Bacillus genomos-
rRNA gene: a rapid tool for identification of Bacillus anthracis. Emerg
pecies, novel members of Bacillus rRNA group 6 isolated from sea
water of the East Sea and the Yellow Sea in Korea. Int J Syst Evol
Siefert, J. L., Larios-Sanz, M., Nakamura, L. K., Slepecky, R. A., Paul,
J. A., Moore, E. R., Fox, G. E. & Jurtshuk, P., Jr (2000). Phylogeny of
Zeigler, D. R. (2003). Gene sequences useful for predicting related-
marine bacillus isolates from the Gulf of Mexico. Curr Microbiol 41,
ness of whole genomes in bacteria. Int J Syst Evol Microbiol 53,
8/2 Ramsay Place, Albury, NSW, 2640 PH: 02 6041 4879 Fax: 02 6041 5783 Trochanteric bursitis is inflammation of the bursa (fluid-filled sac near a joint) at the outside (lateral) point of the hip known as the greater trochanter. When this bursa becomes irritated or inflamed, it causes pain in the hip. This is a common cause of hip pain. Patien
Pfizer Support to European and International Patient Organisations in 2012 The below list contains information on the support Pfizer has provided to European- and international-level patient organisations in 2012, and services contracted from them (e.g. speaker fees). It follows the standards set out in the EFPIA code of practice governing relationships between the pharmaceutical industry
 Strain m4-4T was isolated from surface water samples that
1500 uC for 3 min, increasing at the rate of 40 uC min21 to
were taken and placed in sterile flasks. These were subjected
a final temperature of 3000 uC, which was maintained for
to a shock temperature of 80 uC for 20 min by means of
20 min. Helium was used as carrier gas with a constant
damp heat (Istock et al., 2001). Subsequently, 1 : 100 and
flow of 1 ml min21. Fatty acid methyl esters were identified
1 : 1000 dilutions were made. Aliquots (100 ml) from each
using the mass spectral library search (NIST MS Data Base)
dilution, as well as from the original water samples, were
distributed by the National Institute of Standards and
placed in Petri dishes with marine agar 2216 medium (MA;
Difco) and incubated at 37 uC for 2 days. Cultures were
A combination of Sanger (Nunally, 2005) and 454 Life
purified by subculturing on the same medium and
Sciences sequencing methods (Margulies et al., 2005) were
maintained at –80 uC in 5 % MA and 15 % (w/v) glycerol.
Strain m4-4T was isolated from surface water samples that
1500 uC for 3 min, increasing at the rate of 40 uC min21 to
were taken and placed in sterile flasks. These were subjected
a final temperature of 3000 uC, which was maintained for
to a shock temperature of 80 uC for 20 min by means of
20 min. Helium was used as carrier gas with a constant
damp heat (Istock et al., 2001). Subsequently, 1 : 100 and
flow of 1 ml min21. Fatty acid methyl esters were identified
1 : 1000 dilutions were made. Aliquots (100 ml) from each
using the mass spectral library search (NIST MS Data Base)
dilution, as well as from the original water samples, were
distributed by the National Institute of Standards and
placed in Petri dishes with marine agar 2216 medium (MA;
Difco) and incubated at 37 uC for 2 days. Cultures were
A combination of Sanger (Nunally, 2005) and 454 Life
purified by subculturing on the same medium and
Sciences sequencing methods (Margulies et al., 2005) were
maintained at –80 uC in 5 % MA and 15 % (w/v) glycerol.