Eur Arch Psychiatry Clin Neurosci (1999) 249 : 238–246
I. Leykin · B. Spivak · A. Weizman · I. R. Cohen · M. Shinitzky
Elevated Cellular Immune Response to Human Heat-Shock Protein-60 in Schizophrenic Patients
Received: 26 November 1998 / Accepted: 10 July 1999
Abstract Heat shock protein-60 (HSP60) is implicated
patients and in controls. Titers of IgA against HSP60 were
in several autoimmune diseases as a triggering antigen.
somewhat higher in the sera of schizophrenic patients in
Based on the autoimmune hypothesis of schizophrenia,
comparison to sera of control subjects (P = 0.0605).
we examined cellular and humoral responses againstHSP60 and a series of its peptide fragments with periph-
Key words Autoimmunity · autoantigen · heat shock
eral blood samples of schizophrenic patients and healthy
protein-60 · immune response · schizophrenia
subjects each of group size between 12 to 32 participants. The average stimulation indices of peripheral bloodmononuclear cells (PBMC) to HSP60 were 3.17 ± 0.36
(mean ± SE) for schizophrenic patients and 2.23 ± 0.24(mean ± SE) for healthy subjects, with a significant dif-
A series of immunological abnormalities have been as-
ference between the groups (P = 0.0457). In parallel, 38
cribed to schizophrenia. They include elevated serum im-
synthetic peptide fragments of HSP60, each of 18–21
munoglobulins (Sugerman et al. 1982, DeLisi et al. 1985),
amino acids, were tested for in vitro sensitization of
decreased mitogenic response of peripheral blood lym-
PBMC. With one peptide (p32) the average stimulation
phocytes (Vartanyan et al. 1978, Ganguli et al. 1987,
index of PBMC from schizophrenic patients was signifi-
Chengappa et al. 1995), the presence of morphologically
cantly higher than that obtained for PBMC of control sub-
abnormal large lymphocytes in the blood and bone mar-
jects (P = 0.0006). Comparing the cellular immune re-
row (Vartanyan et al. 1978), increased serum IL-2 recep-
sponse to p32 between patients who were distinctive re-
tor levels (Ganguli and Rabin 1989, Ganguli et al. 1989,
sponders (n = 10) or non-responders (n = 10) to neurolep-
Rapaport et al. 1989, Wilke et al. 1996), decreased IL-2
tic treatment indicated a similar elevation of cellular re-
(Ganguli et al. 1989, Villemain et al. 1989, Ganguli et al.
sponse in these groups. Antibodies against HSP60 were
1992, Ganguli et al. 1995, McAllister et al. 1995) and
screened by dot-blot and ELISA in the sera of the above
IFN-γ production (Rothermundt et al. 1996, Wilke et al.
blood samples. Titers of IgG and IgM against HSP60
1996), and a high serum level of IL-6 (Shintani et al.
were found to be of similar magnitude in schizophrenic
1991). With respect to lymphocyte sub-populations, Ny-land et al. (1980) found a reduced percentage of T cells inschizophrenic patients with acute relapse. Masserini et al. (1990) found an increase of T suppressor lymphocytes in
Acknowledgement This study was supported by a grant from
drug-free schizophrenic patients, while the neuroleptic-
treated patients showed an increase of T helper lympho-cytes. A decrease in the percentage of T cells in schizo-
phrenic patients during acute attack and an increase in the
Department of Biological Chemistry, Weizmann Institute of Science, Rehovot, 76100, Israel
helper/suppressor T cell ratio were found to correlate with
the psychiatric status (Coffey et al. 1983).
As a whole, these findings point to an aberration in the
Ness-Ziona Psychiatric Hospital, Ness-Ziona, Israel
immune function of schizophrenic patients, which couldpromote an autoimmune reaction and, in turn, might con-
A. WeizmanGeha Psychiatric Hospital, Petach-Tikva, Israel
tribute to the psychotic state (Noy et al. 1994). The pres-ence of antibrain autoantibodies in schizophrenic patients
(Heath et al. 1989, Knight et al. 1990, Henneberg et al.
Department of Immunology, Weizmann Institute of Science, Rehovot, 76100, Israel
1994, Yang et al. 1994), which can induce behavioral
changes upon injection to animals (Pandey et al. 1981),
ature and other types of physiological stress (Jindal et al.
supports this possibility. It was therefore proposed that the
1989, Yound 1990). Their precise function is not fully un-
pathogenesis of schizophrenia is associated to some ex-
derstood, but they seem to play an important role as chap-
tent with autoimmune elements (Knight 1984, Wright and
erons (Srivastava 1994). Bacterial HSPs are major im-
Murray 1993, Noy et al. 1994, Wright et al. 1996). In-
mune targets in a broad spectrum of infectious diseases
deed, schizophrenia shares several features with recog-
(Young et al. 1988, Kaufmann 1990, DeNagel and Pierce
nized autoimmune diseases such as Graves’ disease, in-
1993) and have extremely close homologues in mam-
sulin-dependent diabetes mellitus (IDDM), and rheuma-
malian cells (Dudani and Gupta 1989, Winfield 1989). A
toid arthritis (RA) (Knight 1984). Approximately 50%
recent review (Kaufmann 1994) summarizes the evidence
concordance in identical twins was also found in schizo-
for the possibility that HSP molecules may act as trigger-
phrenia (Knight and Adams 1982), as well as an associa-
tion between histocompatibility antigens and disease sus-
If indeed HSPs were to be involved in the pathogene-
ceptibility, with an increase in the frequency of HLA A2,
sis of schizophrenia, then it might be possible to detect
A23 (Amar et al. 1988), A10, A11, A29 (Oscan et al.
their corresponding humoral or cellular responses, which
1996), B7 (Sorokina et al. 1987), and DR2 (Dvorakova et
Indirect support for the autoimmune hypothesis of
schizophrenia is reflected in co-morbidity correlations,
which are typical to autoimmune diseases, and seem toapply to schizophrenia, as well. Finney (1989) reported a
negative correlation between IDDM and schizophrenia,
Thirty two schizophrenic patients of various categories and stages
while in schizophrenic patients the incidence of IDDM is
participated in the study. Demographic distribution of the partici-
much less than in the normal population. A similar nega-
pating patients is given in Table 1. All the patients were free of
tive epidemiological correlation was also observed for
acute or chronic disorders which could affect immune function,
schizophrenia and RA (Mellsop et al. 1974, Ostenberg
and their laboratory blood tests, including CRP and rate of sedi-mentation, were in the normal range. The PANSS psychiatric rat-
1978, Allebeck et al. 1985, Malck-Ahmadi 1985, Spector
ing (Kay et al. 1990) was carried out by an independent group of
and Silman 1987, Spector and Silman 1990, Vinogradov
psychiatrists on the day of blood drawing.
et al. 1991, Eaton et al. 1992). Overall, schizophrenic pa-
Healthy volunteers, mostly of the hospital staff, constituted the
tients have a four- to six-fold reduction in the risk of de-
control group. These were 14 males and 7 females, age 27–61(39.7 ± 11.1 years; mean ± SD).
veloping RA, and even a lower risk for IDDM. Anotherindirect evidence for the assertion of an autoimmune armin schizophrenia comes from our recent reports on psychi-
atric improvements in schizophrenic patients treated withthe immunosuppressive drug azathioprine (Levine et al.
Recombinant human HSP60 was a β-galactosidase fusion con-struct expressed from plasmid pRH710 (Boog et al. 1992). Over-
lapping peptide fragments of human HSP60 p1-p38, each of 18–21
Autoimmunity to heat shock proteins (HSPs) seems to
amino acids, were synthesized and purified as described (Elias et
be involved in the etiology of RA (van Eden et al. 1988,
al. 1991). Recombinant human HSP27, HSP70, HSP90, recombi-
De Graeff-Meeder et al. 1991, Hermann et al. 1991, Life
nant E. coli DnaJ, GrpE, GroEL, GroES, and DnaK were suppliedby StressGen Biotechnologies Corp. (Victoria, Canada). Tetanus
et al. 1991), IDDM (Cohen 1991, Elias et al. 1991, Birk et
Toxoid (TT) was purchased from RIVM (Netherlands).
al. 1996, Maclaren and Atkinson 1997), Behçet’s disease
Peroxidase-conjugated AffiniPure Goat Anti-Human IgG (Fc),
(Pervin et al. 1993, Hasan et al. 1996), and probably in
Alkaline Phosphatase-conjugated AffiniPure Goat Anti-Human IgM
other autoimmune diseases, like multiple sclerosis, ather-
(Fc) and Streptavidin-Peroxidase conjugate were obtained fromJackson ImmunoResearch Laboratories, Inc. (PA, USA). Alkaline
osclerosis, systemic sclerosis, systemic lupus erythemato-
Phosphatase-conjugated Mouse Anti-Human IgA
sus, and Graves’ disease (Rajagopalan et al. 1990, Selmaj
supplied by Pharmingen (USA). Alkaline Phosphatase-conjugated
et al. 1991, Danieli et al. 1992 a, Heufelder et al. 1992, Xu
Rabbit Anti-Human IgE was purchased from DAKO (Denmark).
et al. 1992). However, only scarce information is current-
Biotin Mouse Monoclonal Anti-Human IgG1, IgG2, IgG3, and
ly available on the involvement of HSP induced autoim-munity in schizophrenia. A possible implication of HSPautoimmunity in schizophrenia could be inferred from the
Table 1 Demographic data of the schizophrenia patients of this study
work of Kilidireas et al. (1992), who reported an increaseof antibodies against HSP60 in schizophrenic patients. Re-
cently, Schwarz et al. (1998) reported on elevated anti-HSP60 antibodies in subgroups of 10–20% of schizophren-
ic patients, which correlated with high levels of serum IL-2
and ICAM-1, as well as with increased blood-brain per-
meability. Similarly, Mazeh et al. (1998) have detected an-
tibodies to HSP85 in the serum of schizophrenic patients.
Heat shock proteins are a family of conserved proteins
which are synthesized in response to elevation of temper-
* Conventional neuroleptics, except for clozapine
IgG4 were obtain from Sigma Chemical Co. (USA). Mouse Anti-
night incubation at 4 °C. After washing with phosphate buffered
Human IFN-γ mAb and Biotinylated Anti-Human IFN-γ mAb
saline (PBS)/0.05% Tween 20, the wells were blocked with PBS/
were supplied from Genzyme Diagnostics (USA). Human rIFN-γ
10% fetal calf serum for 2 hours at room temperature. Standards
was purchased from Pharmingen (USA).
and supernatants from the cell cultures were added to the washedwells and incubated for 4 hours at room temperature. Then biotini-lated anti-IFN-γ detecting mAb were added. After an additional in-
Peripheral blood mononuclear cells (PBMC)
cubation for 45 min. at room temperature and washing, the strepta-vidin-peroxidase conjugate was added and incubated for 30 min
Human peripheral blood samples, 10–12 ml with heparin anti-co-
at room temperature and then washed 4 times. A mixture of H2O2
agulant, were drawn in the morning and processed within 4 hours.
and 2,2′-Azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (Sigma
Platelet rich plasma was collected by slow centrifugation (100 g
Chemical Co., USA) was added. The color was developed at room
for 20 min. at room temperature) and discarded. The collected
temperature (10–80 min.) and stopped by adding SDS/N,N-di-
blood cells were suspended in 40 ml of HBSS and layered gently
methyl formamide (Sigma Chemical Co., USA). OD values were
on 10 ml Ficoll-Paque (Pharmacia, Sweden) and centrifugated at
measured at 405 nm and evaluated with the aid of a calibration
900 g for 20 min. at room temperature. The interface layer of pe-
curve. Values of spontaneous IFN-γ secretion, usually below the
ripheral blood mononuclear cells (PBMC), composed predominant-
detection limit, were substracted from the overall reading.
ly of lymphocytes, was collected, and contained 7–9 × 106 cells.
One microliter of different concentrations of HSP (1 µg/µl, 100 ng/µl,
Tests were carried out in 96 microwell plates (Nunk, Denmark) in
10 ng/µl, 1 ng/µl, 100 pg/µl, 10 pg/µl, and 1 pg/µl), human (HSP27,
triplicate in a humidified atmosphere with 5% CO2 at 37 °C. Each
HSP60, HSP70, HSP90) or bacterial (DnaJ, GrpE, GroEL, GroES,
well contained 200,000 PBMC suspended in 200 µl of RPMI 1640
DnaK), diluted in PBS, were applied onto nitrocellulose. The spots
medium supplemented with 2% heat inactivated human AB serum,
were dried and then probed with 200 µl of sera from schizophrenic
2 mM L-glutamine, penicillin (100 units/ml), streptomycin (100 µg/
or control subjects by overnight incubation at room temperature,
ml), gentamycin sulfate (50 µg/ml), 1% non-essential amino acids,
with gentle shaking in 800 µl pH 7.7 buffer containing 60 mM cit-
1 mM sodium pyruvate, 5 × 10–5 M β-mercaptoethanol, and 20
ric acid, 90 mM disodium phosphate, 200 mM sodium chloride,
mM HEPES, pH 7.2. Antigens were added at a final concentration
168 mM sodium hydroxide, and 0.4% Tween 20. After 3 washings
of 10 µg/ml for HSP60 and 25 µg/ml for the peptide fragments of
with the same buffer, positive reactions were detected by HRP
HSP60. Wells with Tetanus Toxoid (TT) (1 µg/ml) served as posi-
goat anti-human IgG-Fc Ab at a final dilution of 1 : 5000 in 200
tive controls. Wells without antigens or mitogens were used as
mM Tris, 200 mM KCl, 0.1% Triton X-100, 10.5 mM phenol, and
2.1 mM CaCl2, pH 8.0, for 4 hours at room temperature with 3
Stimulation of PBMC was scored after 7 days by pulsing with
mg/ml 4-chloronaphthol (Sigma Chemical Co., USA) in methanol
1 µCi per well of 3H-thymidine (Amersham, UK) for the last containing 3% H2O2 as the color releasing substrate. 18 hours. Supernatants were collected and stored at –20 °C andsubsequently used for IFN-γ determination (see below).
The stimulation index (SI) was defined as the ratio of mean
cpm of test cultures divided by the mean cpm of control cultureswithout antigen.
Human HSP60 or TT (as a positive control antigen), diluted inPBS (10 µg/ml), was adsorbed onto microtiterplates by 4 hours in-cubation at room temperature, followed by overnight incubation at
4 °C. After removing unbound protein, the wells were blocked byincubation with PBS/1% BSA for 2 hours at room temperature.
IFN-γ secretion was measured by enzyme immunoassay (Abrams
After 4 washings with PBS/0.05% Tween 20, sera from schizo-
et al. 1992). Microtiter plates (Nunk, Denmark) were coated with
phrenic patients and control subjects, diluted 1/10, were added and
purified mouse antihuman IFN-γ monoclonal antibodies by over-
incubated for 2 hours at 37 °C. Following 4 washings with PBS/
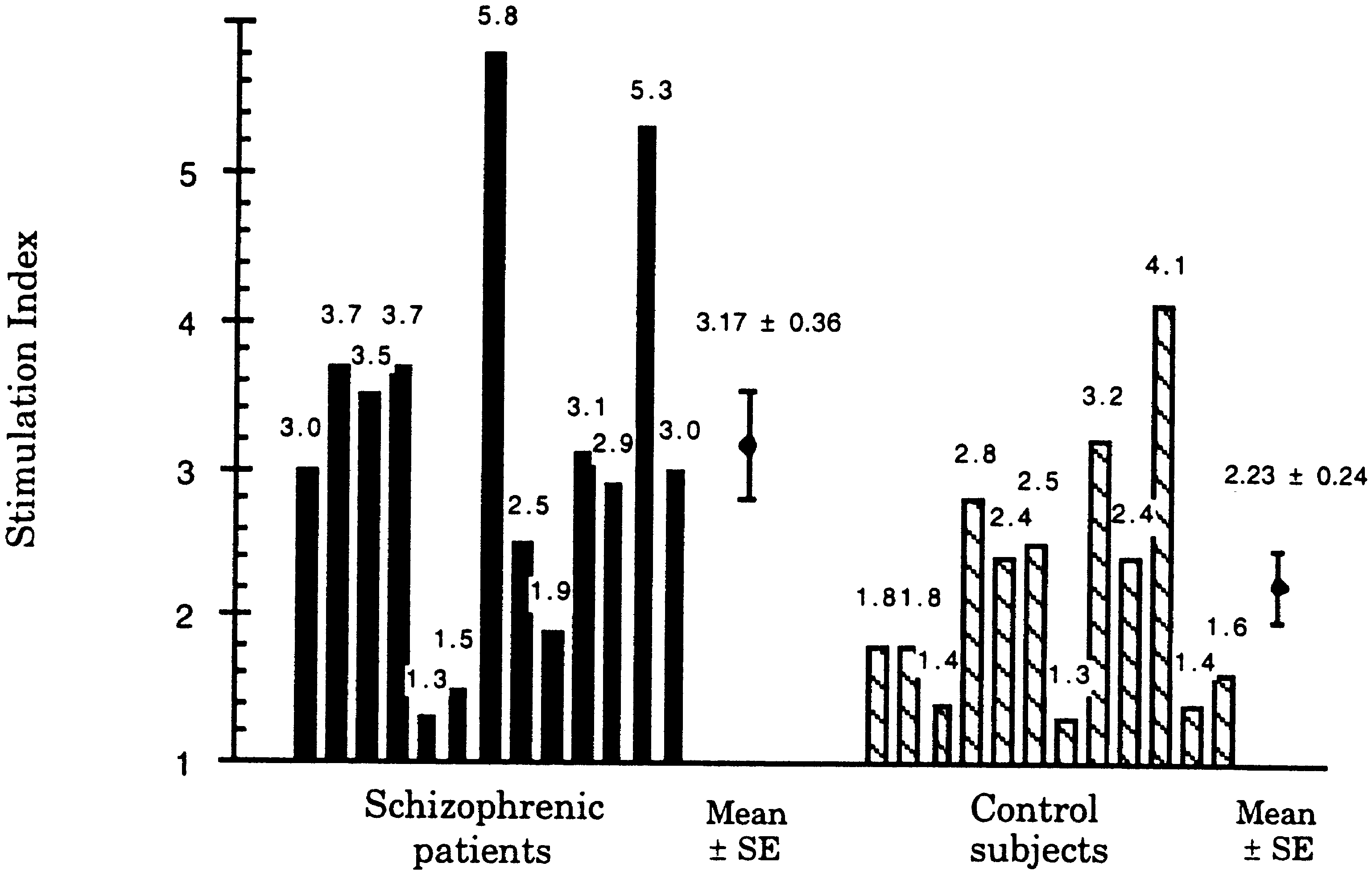
Fig. 1 Stimulation of lympho- cytes from schizophrenic pa- tients (n = 13) and control subjects (n = 12) by human HSP60. The difference in the responses was significant (P = 0.0457)
0.05% Tween 20, peroxidase conjugated anti-human IgG (Fc)
Table 2 Mean Stimulation Index of lymphocytes from schizo-
mAb (1/1000) was added and incubated for 2 hours at room tem-
phrenic patients (n = 8–12) by 25 µg/ml of different fragments of
perature. After washing, a mixture of H2O2 and 2,2′-Azino-bis(3-
HSP60. Indices higher than 2 are marked.
ethylbenzthiazoline-6-sulfonic acid) (Sigma Chemical Co., USA)was added. The color reaction was developed at room temperature
(10–80 min.) and stopped by adding SDS/N,N-dimethyl for-
mamide (Sigma Chemical Co., USA). The plates were scored by
The Mann-Whitney test (two-tailed P value) was used to evaluate
significant differences between experimental and control groups.
This study was carried out in separate segments where
different numbers of subjects participated.
Cell-mediated response against HSP60 was tested by
lymphocyte stimulation in vitro. The results are shown in
Fig. 1. Taking a stimulation index of ≥ 3.0 as positive re-
sponse, the cellular response against HSP60 was positive
in most schizophrenic patients (62%) compared to only
17% in the control group (SI 3.17 ± 0.36 in schizophrenic
patients and 2.23 ± 0.24 in controls, mean ± SE; P =
The above findings prompted us to investigate whether
specific peptides of HSP60 might activate lymphocyte re-
sponses, as has been found in other autoimmune diseases
(Hunt et al. 1993, Anderton et al. 1995, Elias et al. 1995).
Thirty-eight peptide fragments of human HSP60, each of
18–21 amino acids, were assayed for lymphocyte stimula-
tion. Table 2 summarizes the results. One particular frag-
ment (p32) acted as an efficient stimulant of PBMC from
some schizophrenic patients while cells from healthy sub-
jects responded significantly less to this peptide, (3.18 ±
0.38 in schizophrenic patients compared to 1.73 ± 0.10 in
controls, mean ± SE; P = 0.0006; see Fig. 2). An arbitrary
upper normal value of SI ≈ 3 indicates that 17 out of the 33.
32 patients have an abnormal response to p32 (see Fig. 2).
We have further verified whether the spread in response
of PBMC from schizophrenic patients to p32 (see Fig. 2)
is empirically related to the degree of responsiveness to
neuroleptic drugs. As shown in Fig. 3, PBMC from re-
sponders reacted significantly stronger to p32 than PBMCfrom control subjects (P = 0.0147). Non-responders showeda similar higher response as the responders with no signif-icant difference between these groups (P = 0.1655).
Antibody binding to various HSP antigens was assayed
Supernatants from cell cultures stimulated with HSP60
using a dot-blot technique. Sera from schizophrenic pa-
were assayed for IFN-γ. As shown in Fig. 4, IFN-γ secre-
tients, who were either treated with neuroleptics or free of
tion of lymphocytes from schizophrenic patients was sig-
treatment, as well as from normal controls, reacted only
nificantly higher than in control samples (3530 ± 228 pg/
slightly with human HSP27, HSP70, HSP90, and E. coli
ml in schizophrenic patients compared to 2702 ± 301 pg/ml
GrpE, GroEL, GroES, DnaK, and DnaJ. Furthermore, no
in controls, mean ± SE; P = 0.0358). No difference was
difference was detected between sera from schizophrenic
found in IFN-γ secretion, as well as in lymphocyte stimu-
patients and normal subjects, as well as between sera from
lation, between patients treated or untreated with neu-
schizophrenic patients under neuroleptic treatment or neu-
roleptics. Moreover, no correlation was found between the
roleptic-free, in antibody titers to these antigens. Antibody
magnitude of 3H-thymidine incorporation and that of IFN-
titers against HSP60 are summarized in Table 3. The ap-
γ secretion in either the schizophrenic subjects or the con- parent difference between the IgG titers was actually in-trols (data not shown).
significant and was due to exceptional high titers in 2
Fig. 2 Stimulation of lympho- cytes from schizophrenic pa- tients (n = 32) and control sub- jects (n = 21) by peptide p32 of HSP60 (see Table 1). The difference in response between the groups was highly signifi- cant (P = 0.0006). An arbitrary cutoff for the upper normal value was taken as SI ≈ 3. Among the tested patients 53% scored SI above this cutoff Fig. 3 Stimulation of lympho- cytes from responders (n = 10) and non-responders (n = 10) schizophrenic and normal con- trol subjects (n = 10) by pep- tide p32 of HSP60 (see Table 2). The difference in response between responders and con- trol subjects was significant (P = 0.0147), while the differ- ence between responders and non-responders was not signifi- cant (P = 0.1655)
cases in the control group. Subtypes of IgG in the humoral
In general our results on antibodies against HSP60 are
response against HSP60 were also assayed. No difference
in fair agreement with Schwarz et al. (1998) who ob-
was found in the titers of IgG1 between schizophrenics
served that in 80–90% of the schizophrenic patients the
and normal subjects. IgG3 and IgG4 titers were close to
titers fall in the normal range, while in remaining patients
the limit of detection. The IgG2 subtype in schizophrenic
it is significantly elevated. It should be noted that no cor-
sera was slightly lower in comparison to control sera;
relation was observed between the anti HSP antibody
however, this difference was also not significant (data not
titers and the psychiatric evaluation of the individual pa-
tients. No correlation was found between the titers of IgG,
The IgM response to HSP60 appeared to be slightly
IgM or IgA against HSP60 and 3H-thymidine incorpora-
lower in schizophrenic patients (see Table 3), yet with
tion or IFN-γ release shown in Figs. 1–4. Moreover, there
were no differences between the schizophrenic patients
Titers of IgA against HSP60 were found to be some-
and the normal subjects in their antibody and cellular re-
what higher in schizophrenic patients in comparison to the
sponses to TT, except for an IgA response to this antigen
control group. Titers of IgE against HSP60 were below
which was slightly higher in the schizophrenic patients
the detection limit in both schizophrenic patients and
Fig. 4 IFN-γ secretion of lym- phocytes from schizophrenic patients and control subjects after stimulation by human HSP60. The difference in val- ues between the groups was significant (P = 0.0358) Table 3 Titers of antibodies against HSP60 in sera of schizo-
It is reasonable to suppose that organ specific autoim-
phrenic patients (n = 13) and normal subjects (n = 10) presented as
mune diseases should involve some cross reactivity be-
tween a specific fragment of HSP60 and a homologous
epitope on the attacked tissue (Cohen 1991). Specific pep-tide fragments of HSP60 seem to be targeted in IDDM
(Elias et al. 1995), in RA (van Eden et al. 1991, Danieli et
al. 1992 b), and in Behçet’s disease (Stanford et al. 1994,
Direskeneli et al. 1996). The findings described here point
to a specific fragment of HSP60 (p32), which appears to
trigger a T-cell response in PBMC from schizophrenic pa-
tients (see Table 2). Another fragment, p1, elicited a smal-ler response (see Table 2). The responses triggered by theother 36 fragments of HSP60 were similar in magnitudeto those obtained with the PBMC from healthy subjects. The
normal level of immune response to HSP fragments maybe an essential arm in the intricate network of immune ho-
The highly conserved sequence homology between HSPs
meostasis (Kaufmann 1994). Therefore, an autoimmune
of microbial and mammalian origins (Jindal et al. 1989) is
response to a distinct HSP60 peptide could originate ei-
now believed to underlie the etiology of some autoim-
ther from some antigenic cross-reactivity between this
mune responses (van Eden et al. 1988, Elias et al. 1990,
peptide and another target molecule, or from an aberration
Lamb and Young 1990, De Graeff-Meeder et al. 1991,
in the delicate balance of the natural immune response to
Hermann et al. 1991, Res et al. 1991, Danieli et al. 1992 b,
HSP60. On the whole, the present findings provide some
Kaufmann 1994). Accordingly, part of the overt immune
support for the autoimmune hypothesis of schizophrenia
reaction to microbial infection may involve a T-cell re-
(Knight 1984, Noy et al. 1994) and suggest the possible
sponse to the bacterial HSP that can then cross-react with
existence of a specific target epitope, such as peptide p32.
self antigens, which might lead to an autoimmune re-
The putative autoimmune response emanating from
sponse. Evidence from studies on RA (van Eden et al.
HSP60 is at least partially masked by the innate response
1988, De Graeff-Meeder et al. 1991, Hermann et al. 1991,
to HSP60 as a bacterial antigen. This is indicated by the
Life 1991) and Behçet’s disease (Pervin et al. 1993, Hasan
marked overlap between schizophrenic patients and nor-
et al. 1996) supports this concept. In IDDM, experimental
mal subjects in the overt cellular (Fig. 1) and humoral
evidence also suggests such an etiological pattern (Cohen
(Table 3) responses to HSP60. In the cellular responses to
1991, Elias et al. 1991, Birk et al. 1996, Maclaren and
p32 this overlap was reduced considerably which enabled
Atkinson 1997). For other autoimmune diseases like mul-
the assignment of an arbitrary upper value for normal SI
tiple sclerosis, systemic lupus erythematosus, atheroscle-
(see Fig. 2). However, out of the 32 tested patients only 17
rosis, and Graves’ disease, the evidence for the etiological
exhibited SI values above this upper normal value. There-
involvement of HSPs is more indirect (Rajagopalan et al.
fore, at this stage the cellular response to HSP60 or p32
1990, Selmaj et al. 1991, Danieli et al. 1992, Heufelder et
can not provide a reliable tool for biochemical diagnosis
It is interesting that the schizophrenic patients mani-
Birk OS, Elias D, Weiss AS, Rosen A, van der Zee R, Walker MD,
fested increased INF-γ secretion upon HSP60 stimulation
Cohen IR (1996) NOD mouse diabetes: The ubiquitoeus mousehsp60 is a beta-cell antigen of autoimmune T cells. J Autoim-
compared to the controls (see Fig. 4). INF-γ can affect the
neuronal system in several ways. In addition to stimulat-
Boog CJ, E. R. de Greaff-Meeder ER, Lucassen MA, van der Zee
ing differentiation, this cytokine can induce neuronal ex-
R, Voorhorst-Ogink S, van Kooten PJS, Geuze HJ, van Eden
pression of the nitric oxide synthase (NOS) gene (Minc-
W (1992) Two monoclonal antibodies generated against human
Golomb 1996). Cultured neuronal and glial cells were
HSP60 show reactivity with synovial membranes of patientswith juvenile chronic arthritis. J Exp Med 175 : 1805–1810
found to respond to IFN-γ by induction of NOS release
Brunner J, Gattaz WF (1995) Intracerebroventricular injection of
and subsequent cell death (Dawson et al. 1994, Goodwin
phospholipase A2 inhibits apomorphine-induced locomotion in
et al. 1995, Sato et al. 1995). It was also found that IFN-γ
can increase markedly the susceptibility of the human cor-
Chengappa KN, Ganguli R, Yang ZW, Shurin G, Brar JS, Rabin
BS (1995) Impaired mitogen (PHA) responsiveness and in-
tical neurons to infection by coronavirus (Collins 1995).
creased autoantibodies in Caucasian schizophrenic patients
Binding of IFN-γ to its high-affinity receptor induces a
with the HLA B8/DR3 phenotype. Biol Psychiatry 37 : 546–
rapid activation of phospholipase A2 (PLA2) (Ponzoni
and Cornaglia-Ferraris 1993), which could account for the
Coffey CE, Sullivan JL, Rice JR (1983) T lymphocytes in schizo-
observed increase in PLA2 activity in schizophrenia (No-
Cohen IR (1991) Autoimmunity to chaperonins in the pathogene-
ponen et al. 1993, Brunner and Gattaz 1995, Gattaz et al.
sis of arthritis and diabetes. Annu Rev Immunol 9 : 567–589
1995). In neurons, membrane-bound PLA2 plays an es-
Collins AR (1995) Interferon gamma potentiates human coron-
sential role in signal transduction by affecting neurotrans-
avirus OC43 infection of neuronal cells by modulation of HLA
mitter release and receptor sensitivity, with an overall re-
class I expression. Immunol Invest 24 : 977–986
Danieli MG, Candela M, Ricciatti AM, Reginelli R, Danieli G,
duction in neurotransmission (including dopaminergic),
Cohen IR, Gabrielli A (1992 a) Antibodies to mycobacterial
which in turn, may result in dopaminergic alterations with
65kDa heat shock protein in systemic sclerosis (scleroderma). J
schizophrenic symptoms (Brunner and Gattaz 1995, Gat-
Danieli MG, Markovitz D, Gabrielli A, Corvetta A, Giorgi PL,
Van der Zee R, van Embden JD, Danieli G, Cohen IR (1992 b)
We have previously detected abnormal humoral and
Juvenile rheumatoid arthritis patients manifest immune reactiv-
cellular autoimmune responses against autologous throm-
ity to the mycobacterial 65-kDa heat shock protein, to its 180–
bocytes in schizophrenic patients (Deckmann et al. 1996).
188 peptide, and to a partially homologous peptide of the pro-
Of key importance remains the question whether the p32
teoglycan link protein. Clin Immunol Immunopathol 64 : 121–128
peptide of HSP60 cross reacts immunologically with a
Dawson VL, Brahmbhatt HP, Mong JA, Dawson TM (1994) Ex-
specific thrombocyte epitope, which might initiate the au-
pression of inducible nitric oxide synthase causes delayed neu-
toimmune response. This possibility is currently under in-
rotoxicity in primary mixed neuronal-glial cortical cultures.
vestigation in our laboratory. A number of other questions
related to the possible relevance of autoimmunity to the
De Graeff-Meeder ER, Van der Zee R, Rijkers GT, Schuurman HJ,
Kuis W, Bijlsma JWJ, Zegers BJM, van Eden W (1991)
psychopathological features of schizophrenia remain to be
Recognition of human 60 kD heat shock protein by mononu-
investigated. These include the mode of propagation of
clear cells from patients with juvenile chronic arthritis. Lancet
the autoimmune response from the periphery to the CNS
and the identification of the CNS target antigen. Once
Deckmann M, Shinitzky M, Leykin I, Cheng D, Guy J, Avnon M,
Salganik I, Amiri Z, Schlossberg A, Leibu E, Rafael C (1996)
these issues are clarified and further confirmed by inde-
Humoral and cellular response against autologous platelets in
pendent routes, it might be possible to consider immuno-
schizophrenia – clinical and pathophysiological implications.
logical regimens for the treatment of schizophrenia.
Italian J of Psychiatry and Behavioural Sciences 6 : 29–34
DeLisi LE, King AC, Targum S (1985) Serum immunoglobulin
Acknowledgements We thank Dr. Alpha Peled and Mrs. Anna
concentrations in patients admitted to an acute psychiatric in-
Gelfand for their technical assistance. M. S. is the Olin-Sang Pro-
patient service. Br J Psychiatry 145 : 661–665
DeNagel DC, Pierce SK (1993) Heat shock proteins in immune re-
Direskeneli H, Hasan A, Shinnik T, Mizushima Y, van der Zee R,
Fortune F, Stanford MR, Lehner T (1996) Recognition of B-Cell Epitopes of the 65 kDa HSP in Behçet’s Disease. Scand JImmunol 43 : 464–471
Abrams JS, Roncarolo MG, Yssel H, Andersson U, Gleich GJ, Sil-
Dudani AK, Gupta RS (1989) Immunological characterization of a
ver JE (1992) Strategies of anti-cytokine monoclonal antibody
human homolog of the 65-kilodalton mycobacterial antigen.
development: Immunoassay of IL-10 and IL-5 in clinical sam-
Dvorakova M, Majsky A, Zvolsky P (1989) Incidence of HLA-DR
Allebeck P, Rodvall Y, Wistedt B (1985) Incidence of rheumatoid
antigens in endogenous psychoses. Cesk Psychiatr 85 : 217–222
arthritis among patients with schizophrenia, affective psychosis
Eaton WW, Hayward C, Ram R (1992) Schizophrenia and
and neurosis. Acta Psych Scand 71 : 615–619
rheumatoid arthritis: a review. Schizophr Res 6 : 181–192
Amar A, Zohar M, Tiwari J, Brautbar B (1988) HLA and schizo-
Elias D, Markovits D, Reshef T, Van der Zee R, Cohen IR (1990)
phrenia in Israel. Isr J Med Sci 24 : 28–31
Induction and therapy of autoimmune diabetes in the non-
American Psychiatric Association (1994) Diagnostic and Statisti-
obese diabetic (NOD/Lt) mouse by a 65kDa heat shock protein.
cal Manual of Mental Disorders, 4th ed. Washington, DC:
Elias D, Reshef T, Birk OS, van der Zee R, Walker MD, Cohen IR
Anderton SM, van der Zee R, Prakken B, Noordzij A, van Eden W
(1991) Vaccination against autoimmune mouse diabetes using
(1995) Activation of T cells recognizing self 60-kD heat shock
a T cell epitope of human 65kDa heat-shock protein. Proc Natl
protein can protect against experimental arthritis. J Exp Med
Elias D, Markus H, Reshef T, Ablamunits V, Cohen IR (1995) In-
Lamb JR, Young DB (1990) T cell recognition of stress proteins: a
duction of diabetes in standard mice by immunization with the
link between infectious and autoimmune diseases. Mol Biol
p277 of a 60-kDa heat shock protein. Eur J Immunol 25 : 2851–
Levine J, Susnovski M, Handzel ZT, Leykin I, Shinitzky M (1994)
Finney GOH (1989) Juvenile onset diabetes and schizophrenia?
Treatment of schizophrenia with an immunosuppressant. Lan-
Ganguli R, Rabin BS, Kelly RH, Lyte M, Ragu U (1987) Clinical
Levine J, Gutman J, Feraro R, Levy P, Kimhi R, Leykin I (1997)
and laboratory evidence of autoimmunity in acute schizophre-
Side effect profile of azathioprine in the treatment of chronic
schizophrenic patients. Neuropsychobiology 36 : 172–176
Ganguli R, Rabin BS (1989) Increased serum interleukin 2 recep-
Life PF, Bassey EOE, Gaston HJS (1991) T-cell recognition of
tor concentration in schizophrenic and brain-damaged subjects.
bacterial heat shock proteins in inflammatory arthritis. Im-
Ganguli R, Rabin BS, Belle SH (1989) Decreased interleukin-2
Maclaren NK, Atkinson MA (1997) Insulin-dependent diabetes
production in schizophrenic patients. Biol Psychiatry 26 : 427–
mellitus: the hypothesis of molecular mimicri between islet cell
antigens and microorganisms. Mol Med Today 3 : 76–83
Ganguli R, Brar JS, Solomon W, Chengappa KN, Rabin BS (1992)
Malck-Ahmadi P (1985) Rheumatoid arthritis and schizophrenia:
Altered interleukin-2 production in schizophrenia: association
are they mutually exclusive? Semin Arthritis Rheum 15 : 70–72
between clinical state and autoantibody production. Psychiatry
Masserini C, Vita A, Basile R, Morselli R, Boato P, Peruzzi C,
Pugnetti L, Ferrante P, Cazzullo CL (1990) Lymphocyte sub-
Ganguli R, Brar JS, Chengappa KR, DeLeo M, Yang ZW, Shurin
sets in schizophrenic disorders. Relationships with clinical,
G, Rabin BS (1995) Mitogen-stimulated interleukin-2 produc-
neuromorphological and treatment variables. Schizophr Res 3 :
tion in never-medicated, first-episode schizophrenic patients.
The influence of age at onset and negative symptoms. Arch
Mazeh D, Sirota P, Patya M, Novogrotsky A (1998) Antibodies to
neuroblastoma cell line proteins in patients with schizophrenia.
Gattaz WF, Schmitt A, Maras A (1995) Increased platelet phos-
pholipase A2 activity in schizophrenia. Schizophr Res 16 : 1–6
McAllister CG, van Kammen DP, Rehn TJ, Miller AL, Gurklis J,
Goodwin JL, Uemura E, Cunnick JE (1995) Microglial release of
Kelley ME, Yao J, Peters JL (1995) Increases in CSF levels of
nitric oxide by the synergistic action of beta-amyloid and IFN-
interleukin-2 in schizophrenia: effects of recurrence of psy-
chosis and medication status. Am J Psychiatry 152 : 1291–1297
Hasan A, Fortune F, Wilson A, Warr K, Shinnick T, Mizushima Y,
Mellsop GW, Koadlow L, Syme J, Whittingham S (1974) Absence
van der Zee R, Stanford MR, Sanderson J, Lehner T (1996)
of rheumatoid arthritis in schizophrenia. Aust NZJ Med 4 :
Role of γδ T cells in pathogenesis and diagnosis of Behçet’s
Minc-Golomb D, Yadid G, Tsarfati I, Resau JH, Schwartz JP
Heath RG, McCarron KL, O’Neil CE (1989) Antiseptal brain anti-
(1996) In vivo expression of inducible nitric oxide synthase in
body in IgG of schizophrenic patients. Biol Psychiatry 25 :
cerebellar neurons. J Neurochem 66 : 1504–1509
Noponen M, Sanfilipo M, Samanich K, Ryer H, Ko G, Augrist B,
Henneberg AE, Horter S, Ruffert S (1994) Increased prevalence of
Wolkin A, Duncan E, Rotrosen J (1993) Elevated PLA2 activ-
antibrain antibodies in the sera from schizophrenic patients.
ity in schizophrenic and other psychiatric patients. Biol Psychi-
Hermann E, Lohse AW, Van der Zee R, van Eden W, Mayet WJ,
Noy S, Achiron A, Laor N (1994) Schizophrenia and Autoimmu-
Probst P, Poralla T, Meyer zum Büschenfelde KH, Fleisher B
nity – A Possible Etiological Mechanism? Neuropsychobiol-
(1991) Synovial fluid-derived Yersinia-reactive T cells re-
sponding to human 65-kDa heat-shock protein and heat-stressed
Nyland H, Naess A, Lunde H (1980) Lymphocyte subpopulations
antigen-presenting cells. Eur J Immunol 21 : 2139–2143
in peripheral blood from schizophrenic patients. Acta Psychiatr
Heufelder AE, Wenzel BE, Bach RS (1992) Cell surface localiza-
tion of a 72 kilodalton heat shock protein in retroocular fibro-
Oscan ME, Taskin R, Banoglu R, Batacan M, Tuncer E (1996)
blast from patients with Graves’ ophtalmopathy. J Clin En-
HLA Antigens in Schizophrenia and Mood Disorders. Biol
Hunt DW, Corson L, Baker HD, Levy JG, Petty RE (1993) Rela-
Ostenberg E (1978) Schizophrenia and rheumatic disease. A study
tionship between collagen-induced and adjuvant arthritis in the
on the concurrence of inflammatory joint diseases and a review
of 58 case-records. Acta Psych Scand 58 : 339–359
Jindal S, Dudani AK, Singh B, Harley CB, Gupta RS (1989) Pri-
Pandey RS, Gupta AK, Chaturvedi UC (1981) Autoimmune model
mary structure of a human mitochondrial protein homologous
of schizophrenia with special reference to antibrain antibodies.
to the bacterial and plant chaperonins and to the 65-kilodalton
mycobacterial antigen. Mol Cell Biol 9 : 2279–2283
Pervin K, Childerstone A, Shinnik T, Mizushima Y, van der Zee
Kaufmann SHE (1990) Heat shock proteins and the immune re-
R, Hasan A, Vaughan R, Lehner T (1993) T cell epitope ex-
pression of mycobacterial and homologous human 65-kilodal-
Kaufmann SHE (1994) Heat Shock Proteins and Autoimmunity: A
ton heat shock protein peptides in short term cell lines from pa-
Critical Appraisal. Int Arch Allergy Immunol 103 : 317–322
tients with Behçet’s Disease. J Immunol 151 : 2273–2282
Kay SR, Poler LA, Eiszbein A (1990) Positive and negative syn-
Ponzoni M, Cornaglia-Ferraris P (1993) Interferon-gamma-stimu-
drome scale (PANSS). Toronto Multi-Heath System Inc
lated and GTP-binding proteins-mediated phospholipase A2
Kilidireas K, Latov N, Strauss DH, Gorig AD, Hashim GA, Gor-
activation in human neuroblasts. Biochem J 294 : 893–898
man JM, Sadiq SA (1992) Antibodies to the human 60 kDa
Rajagopalan S, Zordan T, Tsokos GC, Datta SK (1990) Pathogenic
heat-shock protein in patients with schizophrenia. Lancet 340 :
anti-DNA autoantibody-inducing T helper cell lines from pa-
tients with active lupus nephritis: Isolation of CD4–8– T helper
Knight JG, Adams DD (1982) The genetic basis of autoimmune
cell lines that express the γ/δ T-cell antigen receptor. Proc Natl
disease. Ciba Foundation Symposium 90 : 35–56
Knight JG (1984) Is Schizophrenia an Autoimmune Disease? – A
Rapaport MH, McAllister CG, Pickar D, Nelson DL, Paul SM
Review. Meth and Find Exp Clin Pharmacol 6 : 394–403
(1989) Elevated levels of soluble interleukin 2 receptors in
Knight JG, Knight A, Menkes DB, Mullen PE (1990) Autoanti-
schizophrenia. Arch Gen Psychiatry 46 : 291–292
bodies against brain septal region antigen specific to unmed-icated schizophrenia? Biol Psychiatry 28 : 467–474
Res PCM, Thole JER, De Vries RRP (1991) Heat shock proteins
Vartanyan ME, Kolyaskina GI, Lozovsky OV, Burbaeva G S, Ig-
in immunopathology. Current Opinion in Immunology 3 : 924–
natov SA (1978) Aspects of humoral and cellular immunity in
schizophrenia. Birth Defects 14 : 339–364
Rothermundt M, Arolt V, Weitzsch C, Eckhoff D, Kirchner H
Villemain F, Chatenoud L, Galinowski A, Homo-Delarche F,
(1996) Production of cytokines in acute schizophrenic psycho-
Ginestet D, Loo H, Zarifian E, Bach JF (1989) Aberrant T cell-
mediated immunity in untreated schizophrenic patients: defi-
Sato I, Kim Y, Himi T, Murota S (1995) Induction of calcium-in-
cient interleukin-2 production. Am J Psychiatry 146 : 609–616
dependent nitric oxide synthase activity in cultured cerebellar
Vinogradov S, Gottesman II, Moises HW, Nicol S (1991) Nega-
granule neurons. Neurosci Lett 184 : 145–148
tive association between schizophrenia and rheumatoid arthri-
Schwarz MJ, Riedel M, Gruber R, Müller N, Ackenheil M (1998)
Autoantibodies against 60-kDa heat shock protein in schizo-
Wilke I, Arolt V, Rothermundt M, Weitzsch C, Hornberg M,
phrenia. Eur Arch Psychiatry Clin Neurosci 248 : 282–288
Kirchner H (1996) Investigations of cytokine production in
Selmaj K, Brosnan CF, Raine CS (1991) Colocalization of Lym-
whole blood cultures of paranoid and residual schizophrenic
phocytes bearing γ/δ T-cell receptor and heat shock protein
patients. Eur Arch Psychiatry Clin Neurosci 246 : 279–284
hsp65+ oligodendrocytes in multiple sclerosis. Proc Natl Acad
Winfield JB (1989) Stress proteins, arthritis, and autoimmunity.
Shintani F, Kanba S, Maruo N, Nakaki T, Nibuya M, Suzuki E,
Wright P, Murray RM (1993) Schizophrenia: prenatal influenza
Kinoshita N, Yagi G (1991) Serum interleukin-6 level in schiz-
ophrenic patients. Life Sci 49 : 661–664
Wright P, Sham PC, Gilvarry CM, Jones PB, Cannon M, Sharma
Sorokina TT, Evsegneev RA, Levin VI, Semenov GV (1987) Fea-
T, Murray RM (1996) Autoimmune diseases in the pedigrees
tures of the distribution of HLA-antigens among patients with
of schizophrenic and control subjects. Schizophr Res 20 : 261–
endogenous psychoses. Zh Nevropatol Psychiatr 87: 885–888
Spector TD, Silman AJ (1987) Does the negative association be-
Xu Q, Dietrich H, Steiner HJ, Gown AM, Schoel B, Mikuz G,
tween rheumatoid arthritis and schizophrenia provide clues to
Kaufmann SHE, Wick G (1992) Induction of arteriosclerosis in
the aetiology of rheumatoid arthritis? Br J Rheumatol 26 : 307–
normocholesterolemic rabbits by immunization with heat
shock protein 65. Arteriosclerosis Thromb 12 : 789–799
Spector TD, Silman AJ (1990) Rheumatoid arthritis, diabetes, and
Yang ZW, Chengappa KN, Shurin G, Brar JS, Rabin BS, Gubbi
AV, Ganguli R (1994) An association between anti-hippocam-
Srivastava PK (1994) Heat shock proteins in immune response to
pal antibody concentration and lymphocyte production of IL-2
cancer: the Fourth Paradigm. Experientia 50 : 1054–1060
in patients with schizophrenia. Psychol Med 24 : 449–455
Stanford MR, Kasp E, Whiston R, Hasan A, Todryk S, Shinnik T,
Young RA (1990) Stress proteins and immunology. Annu Rev Im-
Mizushima Y, Dumonde DC, van der Zee R, Lehner T (1994)
Heat shock protein peptides reactive in patients with Behçet’s
Young D, Lathigra R, Hendrix R, Sweetser D, Young RA (1988)
disease are uveitogenic in Lewis rats. Clin Exp Immunol 97 :
Stress proteins are immune targets in leprosy and tuberculosis.
Sugerman AA, Southern DL, Curran JF (1982) A study of anti-
body levels in alcoholic, depressive and schizophrenic patients. Ann Allergy 48 : 166–171
van Eden W, Thole JE, van der Zee R, Noordzij A, van Embden
JD, Hensen EJ, Cohen IR (1988) Cloning of the mycobacterialepitope recognised by T lymphocytes in adjuvan arthritis. Na-ture 331 : 171–173
you remember to respect your Breezeways must be kept clear-tion. The office staff will be relocated Village Monthly Community Bulletin Board Student Life’s 10th Annual Inter- national Street Festival! Down- on Saturday, April 12th from There will be exhibits from coun-tries all over the world, ethnic crafts, and traditional decorations.
Who’s in Charge? A Role-Playing Activity to Promote Student Understanding of Governance Issues in an Introduction to Education Course Leading the Way in Community College Teacher Education Programs Capitalizing on role playing simulations, this activity provides the opportunity for students the governance process of a local school board and how board decisions impact various stakeholders
 IgG4 were obtain from Sigma Chemical Co. (USA). Mouse Anti-
night incubation at 4 °C. After washing with phosphate buffered
Human IFN-γ mAb and Biotinylated Anti-Human IFN-γ mAb
saline (PBS)/0.05% Tween 20, the wells were blocked with PBS/
were supplied from Genzyme Diagnostics (USA). Human rIFN-γ
10% fetal calf serum for 2 hours at room temperature. Standards
was purchased from Pharmingen (USA).
IgG4 were obtain from Sigma Chemical Co. (USA). Mouse Anti-
night incubation at 4 °C. After washing with phosphate buffered
Human IFN-γ mAb and Biotinylated Anti-Human IFN-γ mAb
saline (PBS)/0.05% Tween 20, the wells were blocked with PBS/
were supplied from Genzyme Diagnostics (USA). Human rIFN-γ
10% fetal calf serum for 2 hours at room temperature. Standards
was purchased from Pharmingen (USA).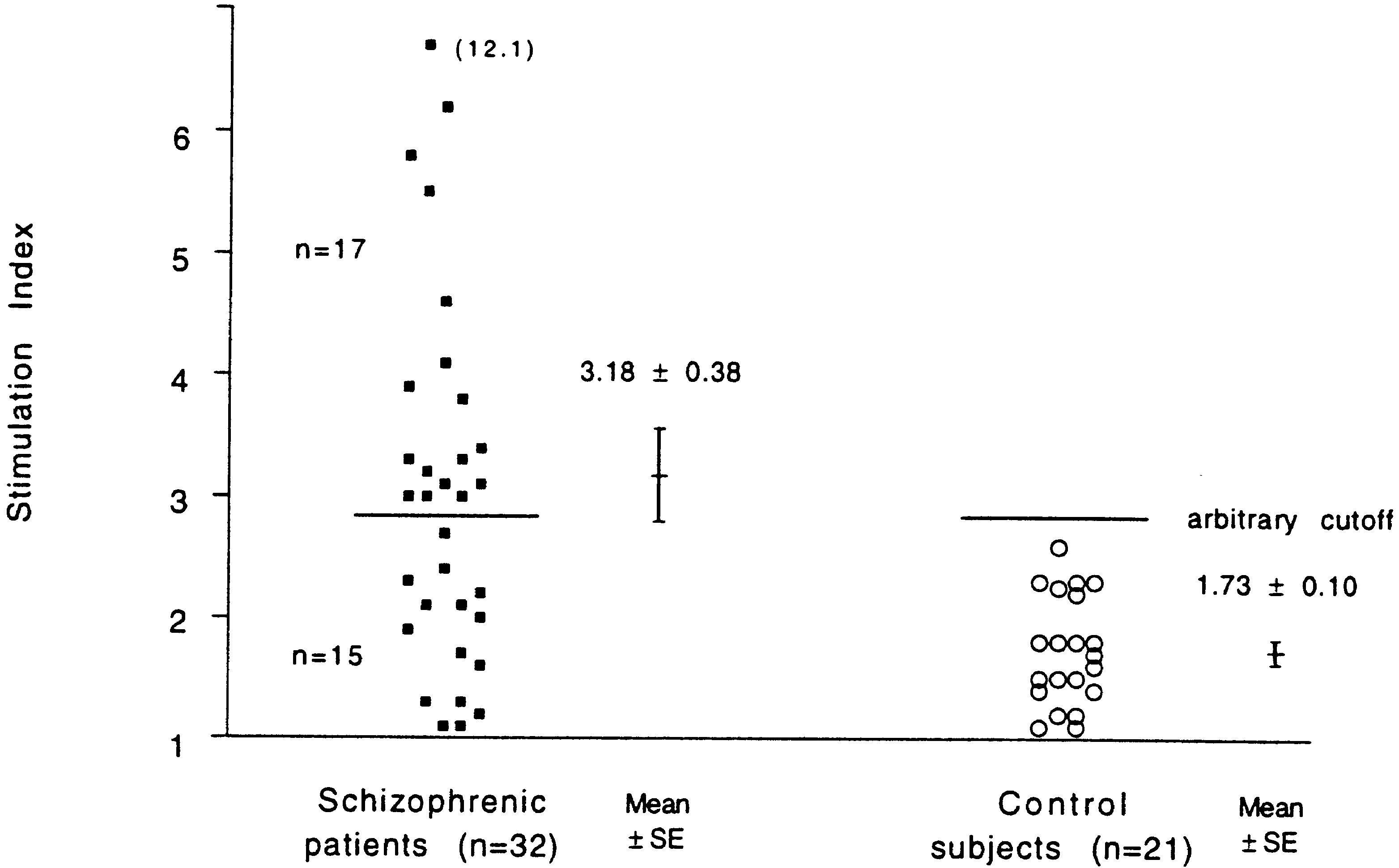
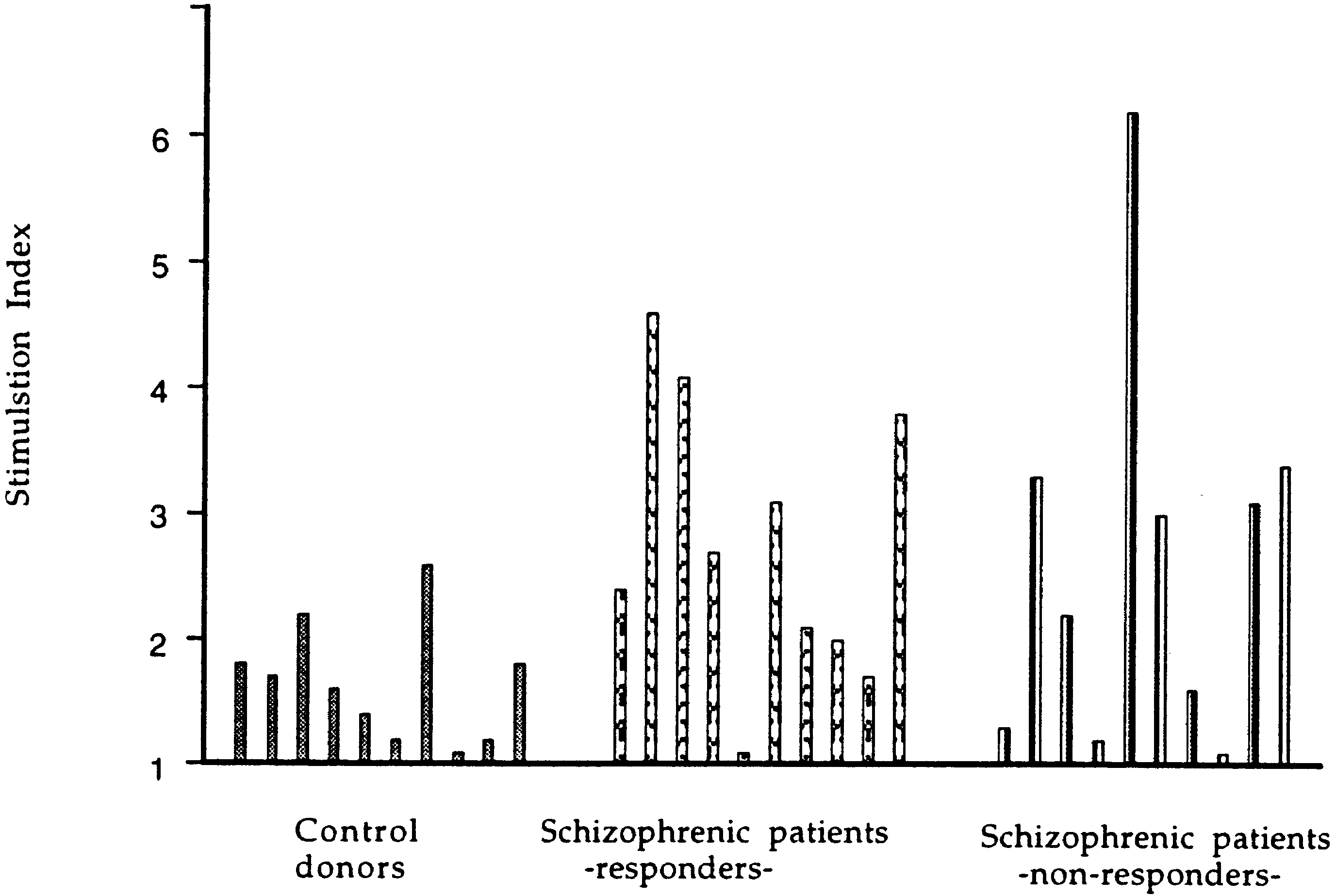 Fig. 2 Stimulation of lympho-
Fig. 2 Stimulation of lympho-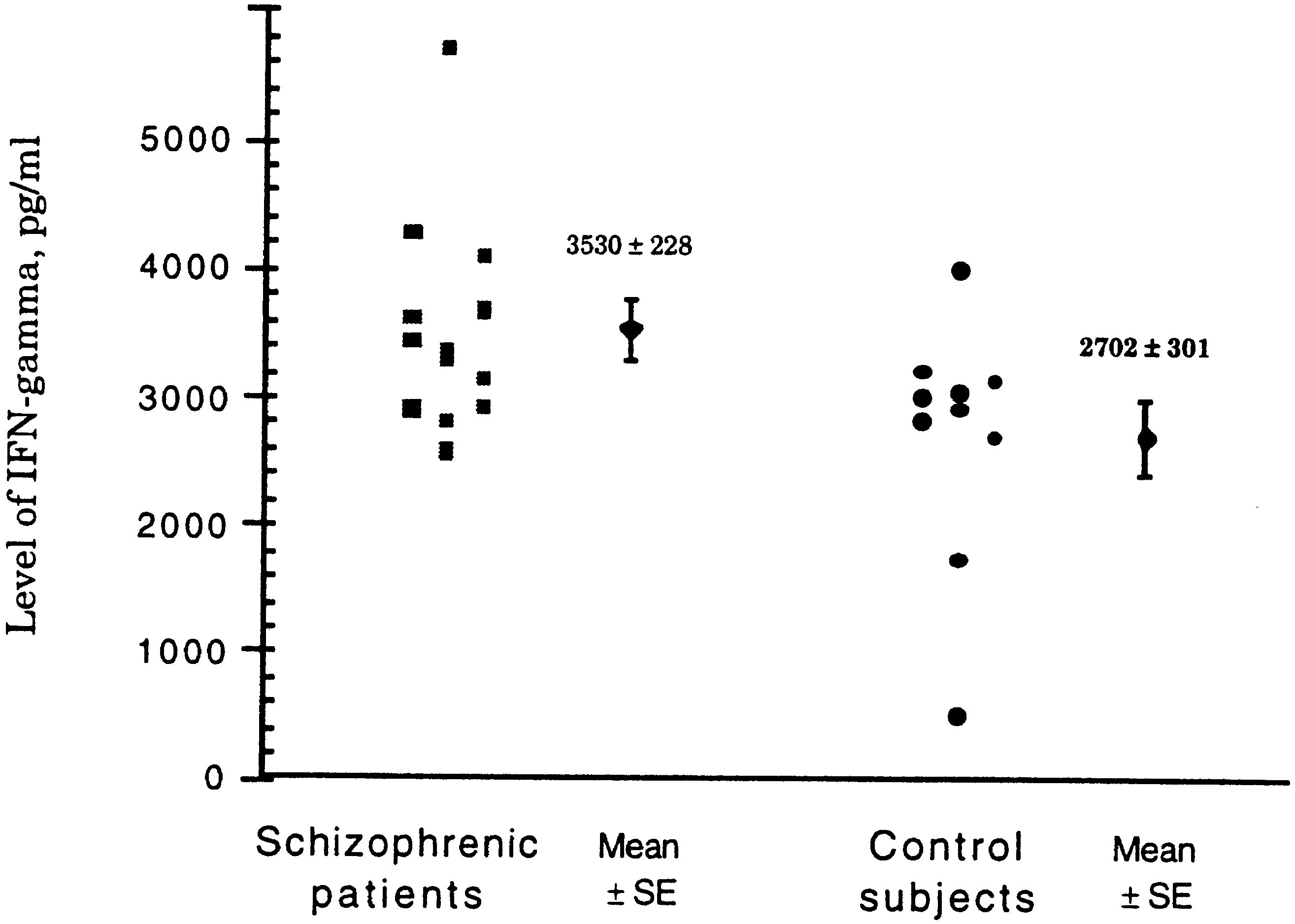 Fig. 4 IFN-γ secretion of lym-
Fig. 4 IFN-γ secretion of lym-