Impaired Th2 Development and Increased Mortality During Schistosoma mansoni Infection in the Absence of CD40/CD154 Interaction1 Andrew S. MacDonald,2 Elisabeth A. Patton,3 Anne C. La Flamme,4 Maria I. Araujo,5 Clive R. Huxtable, Beverley Bauman, and Edward J. Pearce6 The role of CD40/CD154 interaction during infection has primarily focused on pathogens that drive inflammatory Th1 responses. In this study, we show that CD40/CD154 interaction is a fundamental requirement for Th2 response development to the parasitic helminth Schistosoma mansoni. Compared with infected wild-type mice, greatly reduced levels of Th2-associated cytokines were measured both in vitro and in vivo, and no IgE or IgG1 was detected in infected CD154؊/؊ mice. In the absence of an overt Th2 response, no exaggerated Th1 response was mounted by CD154؊/؊ mice. Infected CD154؊/؊ mice suffered severe morbidity and mortality, even though parasitemias in wild-type and CD154؊/؊ mice did not differ significantly. These data indicate that CD40/ CD154 interaction is required to allow development of a Th2-dominated immune response to S. mansoni and support the view that failure to develop such a response can have fatal consequences. The Journal of Immunology, 2002, 168: 4643– 4649.
A memberoftheTNFsuperfamily,CD154(CD40ligand) easesystems(7).Thisapproachhasrevealedthatthereisamarked
is expressed on a wide range of cell types throughout the
heterogeneity in the requirement for CD40/CD154 for appropriate
body that can activate an equally diverse range of CD40-
immune response development depending upon the pathogen in-
expressing cells (1, 2). Interaction of CD40/CD154 provides a ma-
vestigated, with the majority of research in this area focusing on
jor means for T cells to “help” B cells and, through activation of
proinflammatory, Th1-inducing pathogens. However, the outcome
macrophages and dendritic cells (DC)7 to make inflammatory me-
of infection with pathogens that induce Th2 responses in such
diators such as IL-12, also provides a route for skewing the im-
animal models or in individuals with hyperIgM syndrome is much
mune response in a Th1 direction (3). Individuals with hyperIgM
syndrome, an X-linked immunodeficiency caused by a mutation of
The parasitic helminth Schistosoma mansoni is a major public
the CD154 gene, display severely impaired T cell-dependent Ab
health problem on several continents. The immune response to this
responses, with no B cell memory, a reduced Ab repertoire, and
intravascular parasite is strongly Th2 in nature (8), and it is clear
defective germinal center formation (4, 5). As a result, such indi-
that this response provides protection against potentially life-
viduals suffer from recurrent pathogenic microbial infections (6).
threatening aspects of ongoing infection as well as against super-
The development of both CD40- and CD154-deficient mice as
infection (9 –12). However, the mechanisms that control the de-
well as the use of blocking Ab has provided the ability to test therole of CD40/CD154 interaction in a wide range of infectious dis-
velopment of this response, how it is regulated, and how it servesits protective role are still largely unknown.
We have investigated the importance of CD40/CD154 interac-
Department of Microbiology and Immunology, College of Veterinary Medicine, Cor-
tion in the development of an appropriate immune response to S.mansoni by infecting mice that are genetically deficient in CD154.
Received for publication November 16, 2001. Accepted for publication February25, 2002.
Previous work suggested that the CD40/CD154 interaction may be
The costs of publication of this article were defrayed in part by the payment of page
important for Th2 development, because coinjection of S. mansoni
charges. This article must therefore be hereby marked advertisement in accordance
eggs with anti-CD40 Ab results in down-modulation of Th2 de-
with 18 U.S.C. Section 1734 solely to indicate this fact.
velopment (13). Our data have revealed a crucial role for CD154
1 This work was supported by a Wellcome Trust International Prize Traveling Fel-
in Th2 response development during active infection with S. man-
lowship (to A.S.M.) and National Institutes of Health Grant A132573 (to E.J.P.), whois a Burroughs Wellcome Fund Scholar in Molecular Parasitology. Schistosome life
soni. We have found that CD154Ϫ/Ϫ mice infected with S. man-
cycle stages for this work were supplied through Contract NO1-AI55270 from Na-
soni mount a greatly impaired Th2 response and suffer from severe
tional Institute of Allergy and Infectious Disease, National Institutes of Health.
morbidity, exhibiting extreme lung pathology and cachexia that
Current address: Department of Pathobiology, University of Pennsylvania School of
Veterinary Medicine, 3800 Spruce Street, Philadelphia, PA 19104-6008.
eventually lead to death. This sequence of events occurs in the
3 Current address: Department of Comparative Biological Sciences, University of
absence of any overt inflammatory response, suggesting that me-
Wisconsin School of Veterinary Medicine, Madison, WI 53708.
diators such as NO and TNF-␣ are not responsible for the mor-
4 Current address: Victoria University of Wellington School of Biological Sciences,
bidity observed in CD154Ϫ/Ϫ animals.
5 Current address: Servico de Imunologia, Hospital Universitario Professor EdgardSantos, Universidade Federal da Bahia, Bahia, Brazil.
6 Address correspondence and reprint requests to Dr. Edward J. Pearce at the current
Materials and Methods
address: 203D Johnson Pavilion, 3610 Hamilton Walk, University of Pennsylvania,
Philadelphia, PA 19104-6076. E-mail address: [email protected]
CD154Ϫ/Ϫ mice were obtained from The Jackson Laboratory (Bar Harbor,
Abbreviations used in this paper: DC, dendritic cells; MLN, mesenteric lymph node;
SEA, soluble egg Ag; STAg, soluble tachyzoite Ag; WT, wild type.
ME); they were bred in-house and maintained in filter-topped isolator
Copyright 2002 by The American Association of Immunologists
A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
cages, with Bactrim (HiTech Pharmaceutical, Amityville, NY)-supple-
mented water. Wild-type (WT) C57BL/6 mice (Taconic Farms, German-town, NY) were used as controls and similarly maintained on Bactrim-
Expression of surface MHC class II on Mac-1ϩ or B220ϩ cells was quan-
supplemented water. Biomphalaria glabrata snails infected with both
tified by flow cytometry using FITC- or PE-conjugated Ab (I-Ab, B220 or
sexes of S. mansoni and Puerto Rican strain NMRI were obtained from Dr.
Mac-1), purchased from BD PharMingen. Samples were analyzed using a
F. Lewis (Biomedical Research Institute, Rockville, MD). Mice were in-
FACSCaliber flow cytometer and CellQuest software (BD Biosciences,
fected percutaneously with ϳ70 cercariae, weighed regularly, and eutha-
Franklin Lakes, NJ). Proliferation was measured by CFSE (Molecular
nized after significant weight loss (Ͼ20%) was observed in CD154Ϫ/Ϫ
Probes, Eugene, OR) labeling of freshly isolated spleen or MLN cells be-
animals. Adult worms were recovered by perfusion as previously described
fore culture for 5 days with or without 50 g/ml SEA or plate-bound
(14), and samples of lung, liver, and small intestine were collected to quan-
anti-CD3. Cells were then fixed and analyzed as described above.
titate egg deposition (15). Additional samples of lung, liver, and smallintestine were fixed in neutral buffered formalin before staining and his-
tologic examination. Liver granuloma areas were measured on Masson’s
The Student t test was used to determine the statistical significance between
Trichrome-stained sections using Image-Pro Plus software (Media Cyber-
groups. A value of p Ͻ 0.05 was considered a significant difference.
netics, Silver Spring, MD). A minimum of 15 single egg granulomas inclear transverse section were measured per sample. For infection with Tox-oplasma gondii, mice were injected i.p. with 2 ϫ 105 TS4 tachyzoites. Results Morbidity and parasite burden in infected CD154Ϫ/Ϫ mice
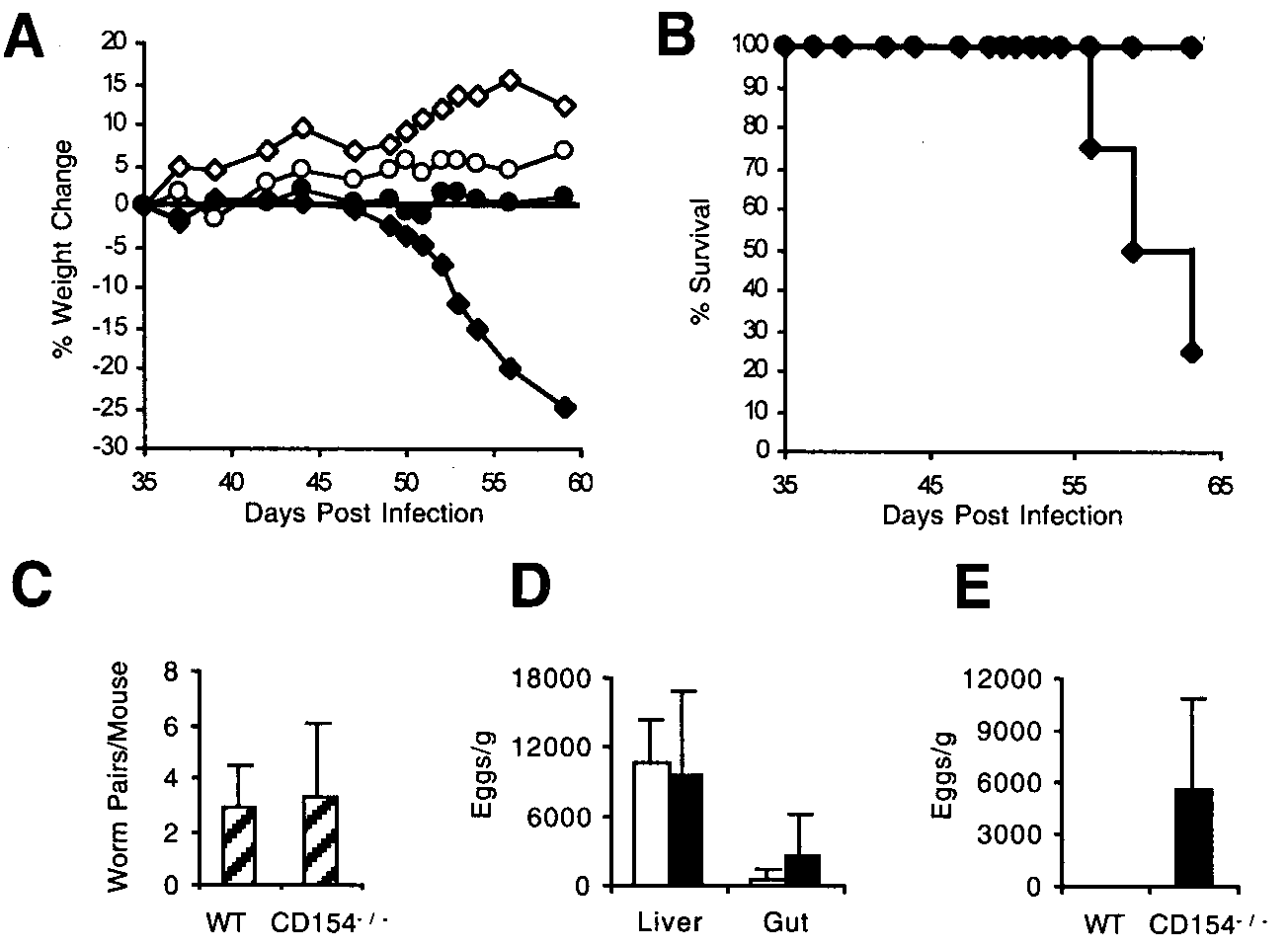
In comparison to infected WT mice, infected CD154Ϫ/Ϫ mice
Soluble egg Ag (SEA) was prepared from isolated schistosome eggs as
showed increasingly severe morbidity from around 7 wk postin-
previously described (16, 17). Soluble tachyzoite Ag (STAg) was prepared
fection (Fig. 1A). Morbidity was characterized by loss of activity,
as described previously (18). SEA (50 g/ml), STAg (20 g/ml), andplate-bound anti-CD3 mAb (BD PharMingen, San Diego, CA; 0.5 g/well)
deterioration of coat condition, deterioration of posture, and
were used to stimulate in vitro cultures.
weight loss. Left unchecked, this eventually culminated in thedeath of infected CD154Ϫ/Ϫ mice within ϳ2 wk of the initiation of
Cell culture and cytokine measurement
weight loss (Fig. 1B). Development of these symptoms was de-
Mesenteric lymph node (MLN) or spleen cells were harvested, and single-
pendent upon patent infection and coincided with the onset of egg
cell suspensions were prepared as previously described (19). Cells were
laying, as CD154Ϫ/Ϫ mice infected with male parasites alone did
counted using trypan blue and resuspended at 107/ml in DMEM (Sigma-
Aldrich, St. Louis, MO) containing 100 U/ml penicillin plus 100 g/mlstreptomycin (Life Technologies, Gaithersburg, MD), 10 mM HEPES (Life
To determine whether cachexia in infected CD154Ϫ/Ϫ mice
Technologies), and 2 mM L-glutamine (Life Technologies). Cells (106/
might be due to differences in parasitemia, we assessed infection
well) were cultured in 96-well flat-bottom plates in 5% CO at 37°C with
intensities in WT and CD154Ϫ/Ϫ animals. However, neither adult
or without the SEA (50 g/ml) or plate-bound anti-CD3 (0.5 g/well).
worm burden (Fig. 1C) nor egg burden in the liver and gut (Fig.
Supernatants were harvested from 24- or 72-h culture and stored at Ϫ20°C
1D) differed significantly between the two groups. In contrast to
for subsequent cytokine analysis by ELISA. Cytokine ELISAs were per-formed on culture supernatants or plasma samples using paired mAb pur-
this, significantly more eggs were recovered from the lungs of
chased from BD PharMingen or purified from hybridoma supernatants in
infected CD154Ϫ/Ϫ mice than from WT mice (Fig. 1E; p Ͻ 0.05).
our laboratory, except for TNF-␣, where the TNF-␣ duoset (R&D Systems,
As might be expected of this chronic infection, weight loss by
Minneapolis, MN) was used. NO production was measured by Greiss re-
CD154Ϫ/Ϫ mice correlated with increasing liver egg burden (not
action (20). Standard ELISPOT protocols were followed to quantitate thenumber of IL-4- and IFN-␥-producing MLN cells, using 0.1 M 2-amino-2
methyl-1-propanol buffer containing 0.6% SeaPlaque agarose and 5-bro-mo-4-chloro-3-indolyl phosphate (1 mg/ml) as a substrate. For peripheral
Pathologic changes in target organs
blood counts, thin blood smears were applied to glass slides, air-dried, and
Examination of gross pathology showed that splenomegaly, al-
fixed in methanol before staining using Hema-3 (Fisher Scientific, Pitts-burgh, PA). The proportion of eosinophils was determined by morphologic
though significant ( p Ͻ 0.05), was less extreme in infected
examination of at least 300 cells/sample in random fields.
CD154Ϫ/Ϫ mice than in infected WT mice (Fig. 2A; p Ͻ 0.02). FIGURE 1.
and CD154Ϫ/Ϫ mice. Cumulative weight change (A)and percent survival (B) of uninfected (open sym-bols) and infected (solid symbols) WT (circles) andCD154Ϫ/Ϫ (diamonds) mice. C, Adult worm burdendid not differ significantly between infected WT orCD154Ϫ/Ϫ mice. Egg burden in liver and gut (D) ofinfected WT (open bars) or CD154Ϫ/Ϫ (solid bars)mice did not differ significantly. However, egg bur-den in the lungs was significantly higher inCD154Ϫ/Ϫ mice (E). Data shown are the mean ϮSD (where shown) for three to eight mice per groupthat were individually assayed and are from one rep-resentative experiment of six performed. FIGURE 2.
Splenomegaly and hepatomegaly in infected WT and
CD154Ϫ/Ϫ mice. Splenomegaly and hepatomegaly were assessed by de-termining spleen (A) and liver (B) size in relation to total body weight foruninfected and infected WT and CD154Ϫ/Ϫ mice. Data shown are themean Ϯ SD of four to eight mice per group that were individually assayedand are from one representative experiment of six performed.
Pronounced hepatomegaly was evident in both groups of mice andwas not significantly different between infected groups (Fig. 2B). More detailed histopathological examination of CD154Ϫ/Ϫ micerevealed several key differences to similarly infected WT mice. Most notably, severe lung pathology was evident in infectedCD154Ϫ/Ϫ animals (Fig. 3). In the lungs, numerous focal granu-lomas were found in CD154Ϫ/Ϫ (Fig. 3D), but not WT (Fig. 3C),mice, coincident with increased numbers of ova and parasite debrisin the lungs of these animals. Although there was extensive in-flammatory change in the lungs of infected WT mice comparedwith uninfected controls, granulomas and parasite debris were rare. Liver and gut pathology was similar in both strains of mice, butfewer granulocytes and mature plasma cells were observed in theabsence of CD154 (not shown). Additionally, liver granulomas
FIGURE 4.
Impaired in vitro Th2 cytokine production by infected
were significantly reduced in size in infected CD154Ϫ/Ϫ mice
CD154Ϫ/Ϫ mice. A, Cytokine production as measured by ELISA in culturesupernatants from MLN cells taken from uninfected (control) or S. man-
compared with WT mice (Fig. 3E; p Ͻ 0.001). Equivalent levels
soni-infected WT or CD154Ϫ/Ϫ mice. MLN cells were then stimulated in
of fibrosis were observed in both groups of infected mice by Mas-
vitro with medium (Ⅺ) or SEA (f) for 72 h before supernatant harvest. B,
son’s Trichrome staining (not shown).
Cytokine production as measured by ELISA in culture supernatants fromspleen cells taken from uninfected (control) or T. gondii-infected WT or
Impaired Th2 responses in infected CD154Ϫ/Ϫ mice
CD154Ϫ/Ϫ mice. Spleen cells were then stimulated in vitro with medium
Adult S. mansoni typically reside within the mesenteric veins of
(Ⅺ) or STAg (o) for 72 h before supernatant harvest. Data shown are the
the murine host. To determine the role of CD40/CD154 interaction
mean Ϯ SD of triplicate wells from three to six mice per group that were
in development of the immune response during helminth infection,
we measured the in vitro production of signature Th2 cytokinesIL-4, IL-5, and IL-13 by MLN cells isolated from infectedCD154Ϫ/Ϫ and WT animals (Fig. 4A). We also assessed in vitro
tures (Fig. 4A). Compared with WT mice, MLN cells from infected
production of the proinflammatory Th1 cytokines, IL-12, TNF-␣,
CD154Ϫ/Ϫ mice made significantly less IL-4, IL-5, and IL-13 in
and IFN-␥, and regulatory mediators, IL-10 and NO, in these cul-
response to SEA than those from infected WT mice ( p Ͻ 0.05). FIGURE 3.
infected WT and CD154Ϫ/Ϫ mice. The histologic appearance of lung tis-sue from uninfected WT (A) orCD154Ϫ/Ϫ (B) mice or infected WT(C) or CD154Ϫ/Ϫ (D) mice. Note themultiple granulomas present only ininfected CD154Ϫ/Ϫ lung tissue (D). Sections were stained with H&E. Original magnification, ϫ20. E, Livergranuloma areas were measured onMasson’s Trichrome-stained sectionsfrom infected WT or CD154Ϫ/Ϫ mice. A minimum of 15 single egg granu-lomas in clear transverse section weremeasured per sample. Data shown arethe mean Ϯ SD of four mice pergroup that were individually assayed.
A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
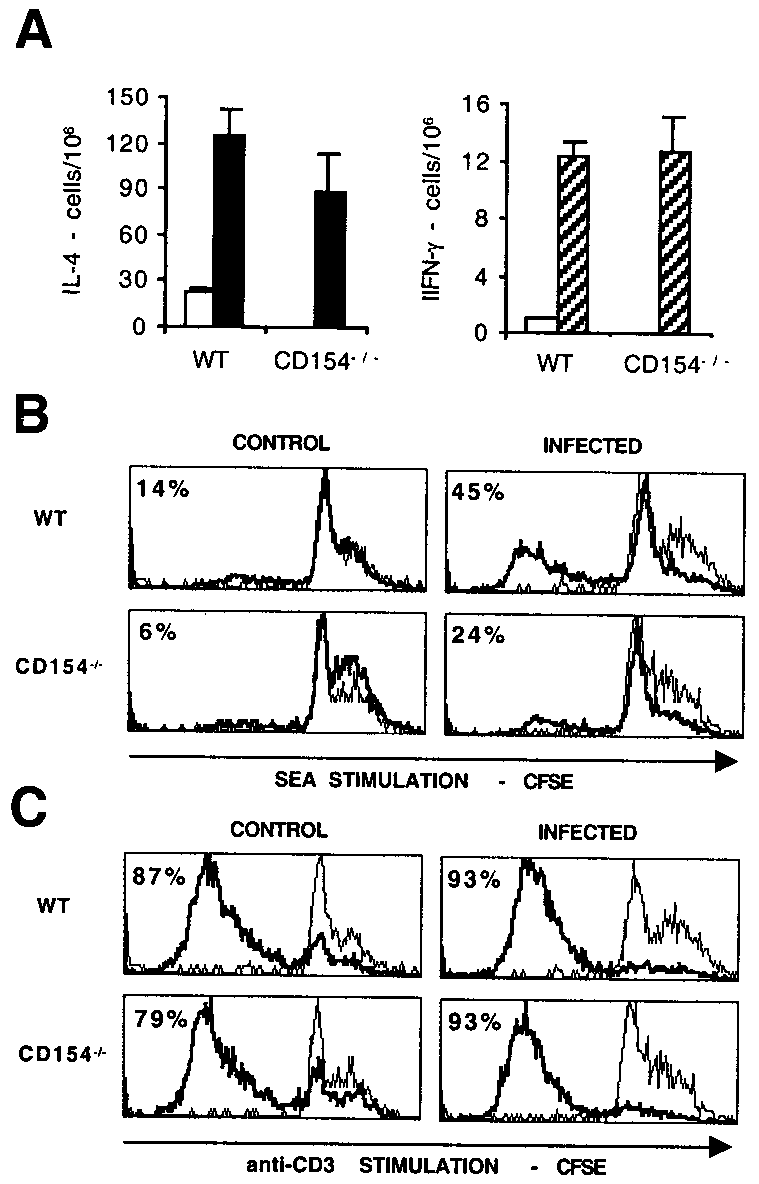
No significant difference in levels of production of IL-12, TNF-␣,or IFN-␥ was seen between the groups, with each of these medi-ators being produced in low amounts. Additionally, we measuredlevels of IL-10 and NO to determine whether the reduced produc-tion of IL-4, IL-5, and IL-13 might be due to enhanced levels ofthese regulatory mediators. However, this proved not to be thecase, with MLN from infected WT mice actually producing sig-nificantly more Ag-specific IL-10 than those from CD154Ϫ/Ϫ mice( p Ͻ 0.01), and with no significant difference between the groupsin the level of NO detected. Consistent with this, immunohisto-chemical staining showed that there was no elevation in iNOSlevels in lung, liver, or gut from infected CD154Ϫ/Ϫ mice com-pared with infected WT mice (not shown). Interestingly, MLNcells from infected CD154Ϫ/Ϫ mice also showed an impaired abil-ity to produce IL-4, IL-5, and IL-13, but not IL-10, IL-12, or NOin response to polyclonal stimulation with plate-bound anti-CD3mAb (not shown). Similar results were obtained from splenocytecultures stimulated with SEA or anti-CD3 (not shown).
Although CD154Ϫ/Ϫ mice infected with S. mansoni appeared to
be unable to mount a Th2 response, the same mice were found tocapably mount a Th1 response to infection with the protozoanparasite Toxoplasma gondii. Parasite-specific IL-12 and IFN-␥were detected in splenocyte cultures from infected CD154Ϫ/Ϫmice at levels that did not differ significantly from those in simi-larly infected WT mice (Fig. 4B).
We reasoned that the impaired Th2 cytokine production seen in
vitro might be due to the reduced initial numbers of cytokine-
FIGURE 5.
Infected CD154Ϫ/Ϫ mice show defective in vitro prolifer-
producing cells or defective cellular proliferation in response to
ative responses. A, ELISPOT was used to quantitate the number of IL-4-
stimulation. To determine whether either of these possibilities might
and IFN-␥-producing MLN cells freshly isolated from uninfected (Ⅺ) or S.
be the case, we examined the ability of MLN cells from WT or
mansoni-infected mice (f or o). Proliferation of WT or CD154Ϫ/Ϫ CFSE-
CD154Ϫ/Ϫ mice to produce IL-4 and IFN-␥ ex vivo by ELISPOT
labeled MLN cells from uninfected (control) or infected mice in response
(Fig. 5A) and to proliferate in response to specific Ag (SEA; Fig.
to 5-day culture with SEA (B) or plate-bound anti-CD3 (C) was deter-
5B) or to a mitogenic stimulus (plate-bound anti-CD3; Fig. 5C)
mined. Histograms represent unstimulated (fine lines) and Ag-stimulated
using CFSE. Although no significant difference was seen in the
(bold lines) lymphocytes, as gated by scatter, and figures represent the
number of freshly isolated MLN cells that produced IL-4 or IFN-␥
percentage of cells that proliferated, based on markers set on unstimulatedsamples.
when obtained from either WT or CD154Ϫ/Ϫ mice (Fig. 5A), thesubsequent ability of these cells to proliferate was somewhat im-paired in CD154Ϫ/Ϫ animals (Fig. 5B). A reduction in the ability
peripheral blood eosinophilia observed in infected CD154Ϫ/Ϫ
of MLN from infected CD154Ϫ/Ϫ mice to proliferate was seen on
mice compared with WT mice (Fig. 6C; p Ͻ 0.001).
stimulation with SEA, but not with anti-CD3, compared with in-
In addition to their defective ability to produce Th2 cytokines in
fected WT mice. However, diminished Th2 cytokine production
vitro and in vivo, infected CD154Ϫ/Ϫ mice displayed impaired Ab
was apparent in cultures exposed to both these stimuli (see above).
production, with disrupted isotype switching (Fig. 7). We mea-
Therefore, impaired proliferation alone cannot account for the re-
sured plasma levels of SEA-specific IgM, IgG1, and IgG2a and
duced ability of infected CD154Ϫ/Ϫ mice to mount a Th2 response
total levels of IgE. No elevated IgE or IgG1, both of which are
in vitro, although it may contribute to the reduced Ag-specific
indicative of a Th2 response in mice, were measured in infected
response to SEA. We have yet to determine whether the cells iso-
CD154Ϫ/Ϫ mice. IgG2a, which is indicative of IFN-␥ production
lated from infected CD154Ϫ/Ϫ mice that proliferate poorly to SEAstimulation are T cells or B cells. The mechanism behind thisdefective Ag-specific proliferation is not yet known, but is unlikelyto be effected by NO or IL-10, because levels of these mediators inculture were not elevated in cells cultured from infectedCD154Ϫ/Ϫ mice compared with WT controls (see above). Impaired Th2 response in infected CD154Ϫ/Ϫ mice in vivo
To ensure that the impaired Th2 response observed in infectedCD154Ϫ/Ϫ mice in vitro was also apparent in vivo, especiallygiven the diminished Ag-specific proliferation we had observed in
FIGURE 6.
Impaired in vivo Th2 responses in infected CD154Ϫ/Ϫ
vitro, we measured plasma levels of IL-4 and IL-5 by ELISA (Fig.
mice. IL-4 (A) and IL-5 (B) production in plasma samples from control
6, A and B). Supporting our in vitro data, levels of both IL-4 and
(uninfected) or S. mansoni-infected WT or CD154Ϫ/Ϫ mice. C, Eosino-
IL-5 were significantly less in infected CD154Ϫ/Ϫ mice than in
philia in peripheral blood from control (uninfected) or S. mansoni-infected
infected WT mice ( p Ͻ 0.01 and p Ͻ 0.03, respectively). A further
WT or CD154Ϫ/Ϫ mice. Data shown are the mean Ϯ SD of five to eight
indication of the impaired ability of CD154Ϫ/Ϫ mice to mount a
mice per group that were individually assayed and are from one experiment
Th2 response in vivo was illustrated by the significantly reduced
IgE, and the reduced numbers of granulocytes evident in their pe-ripheral blood and granulomas. Furthermore, this inability tomount a Th2 response has fatal consequences for infected animals.
In light of the greatly reduced levels of Th2 cytokines in their
plasma and that accumulate in culture supernatants of Ag- oraCD3-stimulated lymphoid organ cells of infected CD154Ϫ/Ϫmice, it is interesting that the frequency of cells that can produceTh2 cytokine is equivalent in infected CD154Ϫ/Ϫ and WT mice asassessed by ELISPOT. This discrepancy may reflect a lower levelof cytokine production by CD154Ϫ/Ϫ animals on a per cell basis. Alternatively, the impaired ability of cells isolated fromCD154Ϫ/Ϫ mice to produce Th2 cytokines after in vitro culturemay be due to a defect in the expansion or survival of respondercells after activation. Supportive of this, we have found that SEA-specific proliferation was reduced in CD154Ϫ/Ϫ animals comparedwith WT, and others have shown that T cell persistence as well aspriming are defective when CD40/CD154 interaction is disrupted
FIGURE 7.
Disrupted isotype switching in infected CD154Ϫ/Ϫ mice.
(21, 22). However, the reduced ability of CD154Ϫ/Ϫ mice to
SEA-specific IgG1, IgG2a, and IgM and total IgE in plasma samples from
mount a Th2 response in vivo, characterized by reduced plasma
uninfected (circles) or S. mansoni-infected (diamonds) WT (open symbols)
IL-4, IL-5, IgG1, and IgE levels and reduced peripheral blood
or CD154Ϫ/Ϫ (solid symbols) mice. Data shown are the mean Ϯ SD ofthree to eight mice per group that were individually assayed.
eosinophilia in these animals, suggests that our in vitro findings arephysiologically relevant.
One interpretation of our data is that B cells play an important
and Th1 response development in mice, was not found in infected
role in Th2 response induction, because in the absence of CD40/
CD154Ϫ/Ϫ animals and was found only at low levels in infected
154 interaction, B cell responses are severely impaired (3). Our
WT animals. Parasite-specific IgM was measured in both groups of
data indicate that B cells are not activated appropriately in
infected mice, but was significantly less in CD154Ϫ/Ϫ than in WT
CD154Ϫ/Ϫ mice during infection, showing reduced activation sta-
tus in terms of MHC class II expression and defective isotypeswitching. Both these outcomes may at least in part be due to the
Impaired MHC class II expression in infected CD154Ϫ/Ϫ mice
reduced levels of IL-4 produced by infected CD154Ϫ/Ϫ mice, be-
In many experimental systems CD40/CD154 has been shown to be
cause IL-4 can act to induce MHC class II up-regulation and fa-
important for optimal activation of B cells (3). One indicator of
cilitate isotype switching to the production of IgE and IgG1 (23).
such activation is the level of MHC class II on the surface of these
Additionally, granuloma cellularity was somewhat different in in-
cells. To determine whether CD40/CD154 interaction was required
fected CD154Ϫ/Ϫ mice, which had a reduced proportion of plasma
for normal B cell activation in our system, we compared the levels
cells compared with WT mice. The role of B cells in schistoso-
of expression of MHC class II on the surface of B cells (B220ϩ)
miasis is intriguing, but as yet not fully resolved. Somewhat con-
and other APC (Mac-1ϩ cells; Table I). B cells and other APC in
flicting reports have suggested that mice lacking B cells either fail
infected CD154Ϫ/Ϫ mice displayed significantly lower levels of
to mount a Th2 response during infection (24) or mount a normal
surface MHC class II than WT mice following infection with S.
Th response but develop more severe disease, failing to regulate
mansoni, although they showed some up-regulation compared with
granulomatous pathology (25). Our data resemble the outcome of
Th2 response, but also exhibit impaired granuloma formation and
Discussion
severe pathology. The emerging role of specific B cell subsets that
Taken together our data provide compelling evidence that CD40/
secrete specific cytokines in immune response polarization (26)
CD154 interaction is necessary to allow appropriate immune re-
raises the interesting possibility that a defective B cell response
sponse development during infection with the parasitic helminth S.
may also directly influence the resultant cytokine profile during
mansoni. This requirement is most evident in the significantly im-
schistosome infection, a possibility that remains to be investigated.
paired ability of these animals to produce Th2 cytokines in vitro
DC represent another CD40-bearing cell type that can be acti-
and in vivo, their inability to make Th2-associated Ab IgG1 and
vated through ligation by CD154 (3, 27, 28). It is possible thatdefective DC function could contribute to the impaired Th2 de-velopment we have observed in infected CD154Ϫ/Ϫ mice. Sup-portive of this hypothesis, we have shown that CD40-deficient,
Table I. MHC class II expression on Mac-1ϩ and B220ϩ cellsa
bone marrow-derived DC exhibit a diminished ability to induceTh2 responses in vivo (29). We have yet to investigate the acti-
vation status of DC isolated from S. mansoni infection in the ab-
sence of CD154. An additional possibility that remains to be ad-dressed is whether signaling to the T cell via CD154 might directly
drive Th2 development during infection (30 –32).
Costimulation has previously been shown to be important for
the development of a Th2 response to S. mansoni infection, as
mice doubly deficient for both CD80 and CD86 fail to mount a
Th2 response to the parasite (33). However, in this case the T cell
a Mean fluorescence intensity of staining for MHC class II on spleen or MLN cells
cytokine profile was reversed, with CD80/86-deficient mice se-
from control (uninfected) or infected WT or CD154Ϫ/Ϫ mice, gated for expression ofMac-1 or B220. ND, Too few cells present to accurately assess this parameter.
creting higher levels of IFN-␥ than WT mice coincident with the
A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
loss of IL-4 production. This contrasts what we have seen during
Acknowledgments
infection of CD154Ϫ/Ϫ mice, where no such compensatory Th1
We thank Eric Y. Denkers for helpful discussion, and Jim Marsh and Kevin
response is noted on loss of the Th2 response. Nevertheless, our
Fitch for use of their imaging system.
data suggest that one outcome of CD40/CD154 interaction mightbe to initiate events that lead to up-regulation of costimulatorymolecules such as CD80 and CD86, although the low levels of
References
these molecules measured during infection of even WT mice has
1. Grewal, I. S., and R. A. Flavell. 1998. CD40 and CD154 in cell-mediated im-
made it difficult to show this experimentally (unpublished obser-
munity. Annu. Rev. Immunol. 16:111.
vations). Nevertheless, it seems that the sequence of events that
2. van Kooten, C., and J. Banchereau. 2000. CD40-CD40 ligand. J. Leukocyte Biol.
leads to Th2 response development originates with and is depen-
dent upon CD40 and CD154 interaction.
3. van Kooten, C., and J. Banchereau. 1997. Functions of CD40 on B cells, dendritic
cells and other cells. Curr. Opin. Immunol. 9:330.
The rapid severe weight loss seen in infected CD154Ϫ/Ϫ mice
4. Callard, R. E., R. J. Armitage, W. C. Fanslow, and M. K. Spriggs. 1993. CD40
bears similarity to the outcome of infection of IL-4Ϫ/Ϫ mice in-
ligand and its role in X-linked hyper-IgM syndrome. Immunol. Today 14:559.
fected with S. mansoni (19). In the case of IL-4Ϫ/Ϫ mice, however,
5. Ramesh, N., R. Fuleihan, and R. Geha. 1994. Molecular pathology of X-linked
in the absence of Th2 response development an inflammatory re-
immunoglobulin deficiency with normal or elevated IgM (HIGMX-1). Immunol. Rev. 138:87.
sponse is observed, characterized by elevated production of in-
6. Notarangelo, L. D., M. Duse, and A. G. Ugazio. 1992. Immunodeficiency with
flammatory mediators such as NO and IFN-␥. This contrasts with
hyper-IgM (HIM). Immunodefic. Rev. 3:101.
what we have noted in infected CD154Ϫ/Ϫ animals, in which no
7. Grewal, I. S., P. Borrow, E. G. Pamer, M. B. Oldstone, and R. A. Flavell. 1997.
such elevated inflammatory response is seen. It is difficult to ad-
The CD40-CD154 system in anti-infective host defense. Curr. Opin. Immunol.
dress whether the absence of CD154 might impair Th1 response
development to S. mansoni, because infection does not induce a
8. Pearce, E. J., P. Casper, J.-M. Grzych, F. A. Lewis, and A. Sher. 1991. Down-
regulation of Th1 cytokine production accompanies induction of Th2 responses
marked Th1 response even in WT mice. Thus, we cannot discount
by a parasitic helminth: Schistosoma mansoni. J. Exp. Med. 173:159.
the possibility that infected CD154Ϫ/Ϫ animals may also mount a
9. Brunet, L. R., D. W. Dunne, and E. J. Pearce. 1998. Cytokine interaction and
defective Th1 response to the parasite.
immune responses during Schistosoma mansoni infection. Parasitology Today14:422.
An additional unexpected finding in infected CD154Ϫ/Ϫ mice
10. Brunet, L. R., M. A. Kopf, and E. J. Pearce. 1999. Schistosoma mansoni: IL-4 is
was the presence of large numbers of eggs and severe pathologic
necessary for concomitant immunity in mice. J. Parasitol. 85:734.
changes in the lungs. This occurred before wk 8 of infection at a
11. Fallon, P. G., E. J. Richardson, G. J. McKenzie, and A. N. McKenzie. 2000.
time when it is rare to find eggs in the lungs of WT mice. Shunting
Schistosome infection of transgenic mice defines distinct and contrasting patho-
to the lungs reflects the development of portal hypertension and the
genic roles for IL-4 and IL-13: IL-13 is a profibrotic agent. J. Immunol. 164:2585.
formation of varices that allow blood to leave the portal system
12. Hoffmann, K. F., A. W. Cheever, and T. A. Wynn. 2000. IL-10 and the dangers
of immune polarization: excessive type 1 and type 2 cytokine responses induce
without passing through the liver. Eggs produced by the parasites
distinct forms of lethal immunopathology in murine schistosomiasis. J. Immunol.
in the portal system are thus able to pass through the venous sys-
tem to the heart and be pumped on to the lungs where they become
13. Martin, D. L., C. L. King, E. Pearlman, E. Strine, and F. P. Heinzel. 2000. IFN-␥
trapped in the capillaries. This indication that infected CD154Ϫ/Ϫ
is necessary but not sufficient for anti-CD40 Ab-mediated inhibition of the Th2response to Schistosoma mansoni eggs. J. Immunol. 164:779.
mice develop severe portal hypertension shortly after egg produc-
14. Smithers, S. R., and R. J. Terry. 1965. The infection of laboratory hosts with
tion begins suggests that there is a failure of vascular regulation in
cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasi-
these animals. Infected WT mice exhibit low, but detectable, levels
of iNOS in and around granulomas in the liver, gut, and lungs, but
15. Cheever, A. W. 1970. Relative resistance of the eggs of human schistosomes to
digestion in potassium hydroxide. Bull. W.H.O. 43:601.
this is not seen in infected CD154Ϫ/Ϫ mice. Therefore, it is pos-
16. Boros, D. L., R. Tomford, and K. S. Warren. 1977. Induction of granulomatous
sible that reduced production of mediators such as NO in this con-
and elicitation of cutaneous sensitivity by partially purified SEA of Schistosoma
text might result in defective vascular regulation, perhaps due to
mansoni. J. Immunol. 118:373.
impaired vasodilation, that could, in turn, lead to more rapid de-
17. Dalton, J. P., S. R. Day, A. C. Drew, and P. J. Brindley. 1997. A method for the
velopment of portal hypertension and its detrimental sequelae.
isolation of schistosome eggs and miracidia free of contaminating host tissues. Parasitology 115:29.
It has previously been shown that IL-4 and IL-13 play important
18. Bliss, S. K., Y. Zhang, and E. Y. Denkers. 1999. Murine neutrophil stimulation
roles in granuloma formation and development of fibrosis during
by Toxoplasma gondii antigen drives high level production of IFN-␥-independent
schistosome infection (11, 34). Interestingly, even in the face of a
severely impaired Th2 response and a virtual absence of produc-
19. Brunet, L. R., F. D. Finkelman, A. W. Cheever, M. A. Kopf, and E. J. Pearce.
1997. IL-4 protects against TNF-␣-mediated cachexia and death during acute
tion of IL-4 and IL-13 to the parasite, infected CD154Ϫ/Ϫ mice did
schistosomiasis. J. Immunol. 159:777.
not show significantly different levels of fibrosis from those in WT
20. Green, L. C., D. A. Wagner, J. Glogowski, P. L. Skipper, J. S. Wishnok, and
mice. It is possible that the very low levels of these mediators
S. R. Tannenbaum. 1982. Analysis of nitrate, nitrite, and [15N]nitrate in biolog-
measured in vitro might be sufficient to allow the development of
ical fluids. Anal. Biochem. 126:131.
fibrosis in vivo, or that other mediators are involved. Moreover, the
21. Grewal, I. S., J. Xu, and R. A. Flavell. 1995. Impairment of antigen-specific
T-cell priming in mice lacking CD40 ligand. Nature 378:617.
fact that a granulomatous response was mounted by CD154Ϫ/Ϫ
22. Miga, A. J., S. R. Masters, B. G. Durell, M. Gonzalez, M. K. Jenkins,
mice indicates that there is a CD4 T cell component in the response
C. Maliszewski, H. Kikutani, W. F. Wade, and R. J. Noelle. 2001. Dendritic cell
longevity and T cell persistence is controlled by CD154-CD40 interactions. Eur.
In summary, our data show that CD40/CD154 interaction is re-
quired to allow Th2 response development to the parasitic hel-
23. Chomarat, P., and J. Banchereau. 1997. An update on interleukin-4 and its re-
ceptor. Eur. Cytokine Network 8:333.
minth S. mansoni, and that the absence of this response has fatal
24. Hernandez, H. J., Y. Wang, and M. J. Stadecker. 1997. In infection with Schis-
consequences for the host. These novel observations have impli-
tosoma mansoni, B cells are required for T helper type 2 cell responses but not
cations for understanding the underlying mechanisms of Th2 re-
for granuloma formation. J. Immunol. 158:4832.
sponse initiation, highlighting the pathophysiological relevance of
25. Jankovic, D., A. W. Cheever, M. C. Kullberg, T. A. Wynn, G. Yap, P. Caspar,
F. A. Lewis, R. Clynes, J. V. Ravetch, and A. Sher. 1998. CD4ϩ T cell-mediated
this receptor/ligand pair in this process, and may impact the de-
granulomatous pathology in schistosomiasis is downregulated by a B cell-depen-
velopment of future immunotherapeutic strategies for other Th2-
dent mechanism requiring Fc receptor signaling. J. Exp. Med. 187:619.
26. Harris, D. P., L. Haynes, P. C. Sayles, D. K. Duso, S. M. Eaton, N. M. Lepak,
L. L. Johnson, S. L. Swain, and F. E. Lund. 2000. Reciprocal regulation of
cytokines and synergy with interleukin-12 for interferon-␥ production. Eur. J. Im-
polarized cytokine production by effector B and T cells. Nat. Immunol. 1:475.
27. Caux, C., C. Massacrier, B. Vanbervliet, B. Dubois, C. Van Kooten, I. Durand,
32. Poudrier, J., D. van Essen, S. Morales-Alcelay, T. Leanderson, S. Bergthorsdottir,
and J. Banchereau. 1994. Activation of human dendritic cells through CD40
and D. Gray. 1998. CD40 ligand signals optimize T helper cell cytokine produc-
cross-linking. J. Exp. Med. 180:1263.
tion: role in Th2 development and induction of germinal centers. Eur. J. Immunol.
28. Cella, M., D. Scheidegger, K. Palmer-Lehmann, P. Lane, A. Lanzavecchia, and
G. Alber. 1996. Ligation of CD40 on dendritic cells triggers production of high
33. Hernandez, H. J., A. H. Sharpe, and M. J. Stadecker. 1999. Experimental murine
levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via
schistosomiasis in the absence of B7 costimulatory molecules: reversal of elicited
APC activation. J. Exp. Med. 184:747.
T cell cytokine profile and partial inhibition of egg granuloma formation. J. Im-
29. MacDonald, A. S., A. D. Straw, N. M. Dalton, and E. J. Pearce. 2002. Cutting
edge: Th2 response induction by dendritic cells: a role for CD40. J. Immunol. 168:537.
34. Chiaramonte, M. G., D. D. Donaldson, A. W. Cheever, and T. A. Wynn. 1999.
30. Blotta, M. H., J. D. Marshall, R. H. DeKruyff, and D. T. Umetsu. 1996. Cross-
An IL-13 inhibitor blocks the development of hepatic fibrosis during a T-helper
linking of the CD40 ligand on human CD4ϩ T lymphocytes generates a costimu-
type 2-dominated inflammatory response. J. Clin. Invest. 104:777.
latory signal that up-regulates IL-4 synthesis. J. Immunol. 156:3133.
35. Mathew, R. C., and D. L. Boros. 1986. Anti-L3T4 Ab treatment suppresses he-
31. Peng, X., A. Kasran, P. A. Warmerdam, M. de Boer, and J. L. Ceuppens. 1996.
patic granuloma formation and abrogates antigen-induced interleukin-2 produc-
Accessory signaling by CD40 for T cell activation: induction of Th1 and Th2
tion in Schistosoma mansoni infection. Infect. Immun. 54:820.
1. Pollock A, Baer G, Pomeroy V, Langhorne P. Physiotherapy treatment ap- 10. Khadilkar A, Phillips K, Jean N, Lamothe C, Milne S, Sarnecka J. Ottawa Panel proaches for the recovery of postural control and lower limb function followingevidence-based clinical practice guidelines for post-stroke rehabilitation. Top Stroke stroke. Cochrane Database Syst Rev . 2003;(2):CD001920. 2. Pomero
Renovierungs-Dampfbremse DIFFLEX RENOVA Technisches Merkblatt Beschreibung/Anwendung Renovierungsdampfbremse für den Einbau von innen oder außen. Schützt die Isoliermaterialien zuverlässig vor Feuchte und Zugluft. Technische Daten Eigenschaft Prüfung monolithische Spezialfunktionsmembran ein Diffusionsäquivalente Luftschichtdicke: Widerstand gegen Wasse
 A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
cages, with Bactrim (HiTech Pharmaceutical, Amityville, NY)-supple-
mented water. Wild-type (WT) C57BL/6 mice (Taconic Farms, German-town, NY) were used as controls and similarly maintained on Bactrim-
Expression of surface MHC class II on Mac-1ϩ or B220ϩ cells was quan-
supplemented water. Biomphalaria glabrata snails infected with both
tified by flow cytometry using FITC- or PE-conjugated Ab (I-Ab, B220 or
sexes of S. mansoni and Puerto Rican strain NMRI were obtained from Dr.
A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
cages, with Bactrim (HiTech Pharmaceutical, Amityville, NY)-supple-
mented water. Wild-type (WT) C57BL/6 mice (Taconic Farms, German-town, NY) were used as controls and similarly maintained on Bactrim-
Expression of surface MHC class II on Mac-1ϩ or B220ϩ cells was quan-
supplemented water. Biomphalaria glabrata snails infected with both
tified by flow cytometry using FITC- or PE-conjugated Ab (I-Ab, B220 or
sexes of S. mansoni and Puerto Rican strain NMRI were obtained from Dr.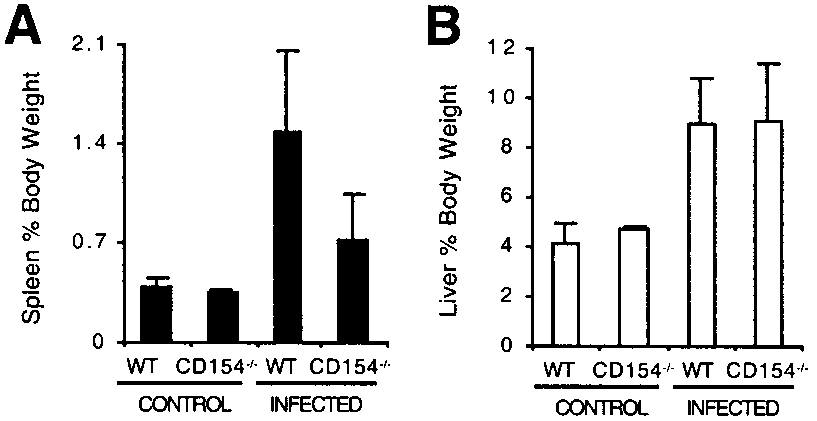
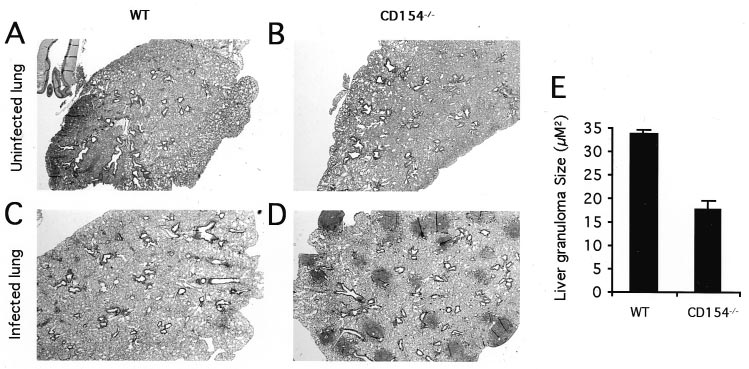
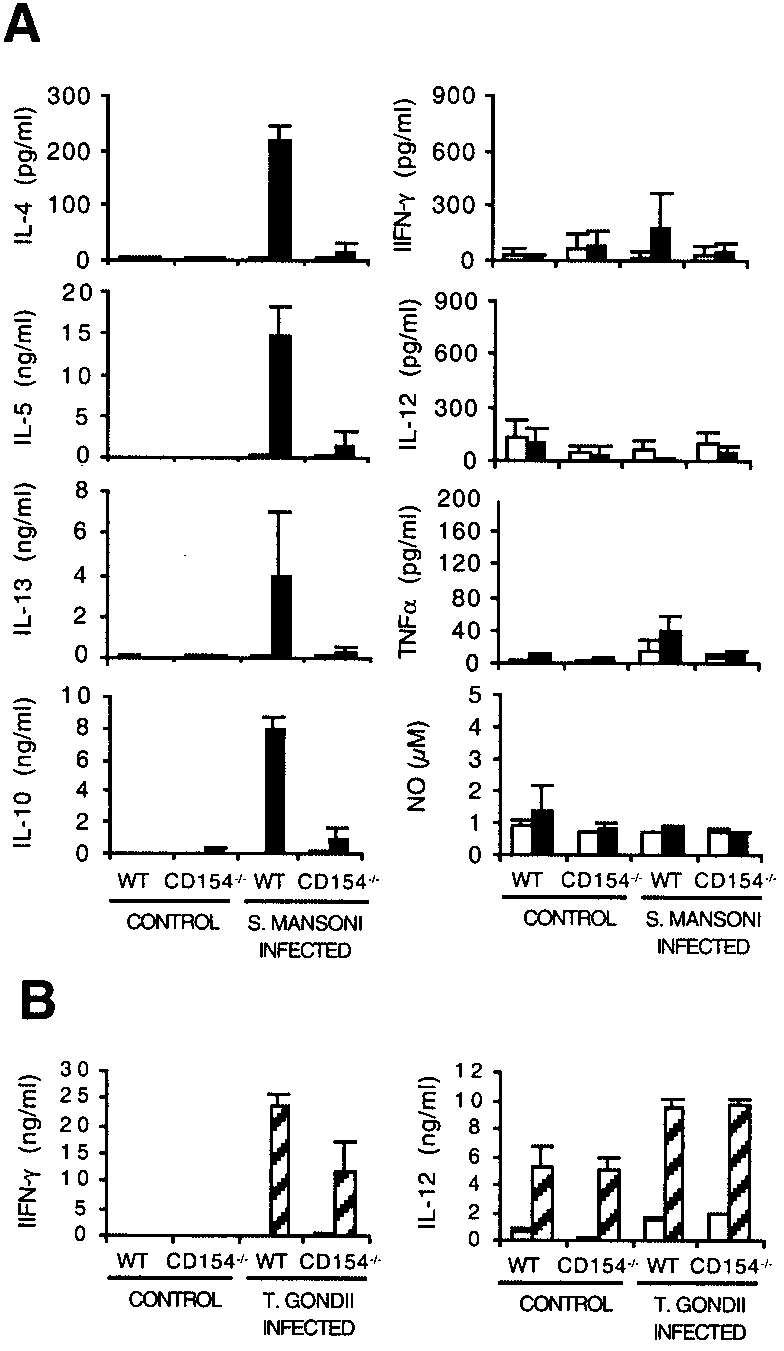 FIGURE 2.
FIGURE 2.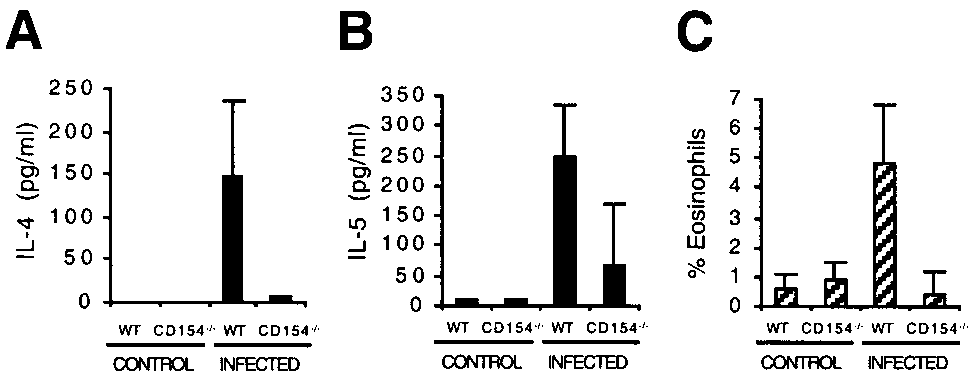
 A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
No significant difference in levels of production of IL-12, TNF-␣,or IFN-␥ was seen between the groups, with each of these medi-ators being produced in low amounts. Additionally, we measuredlevels of IL-10 and NO to determine whether the reduced produc-tion of IL-4, IL-5, and IL-13 might be due to enhanced levels ofthese regulatory mediators. However, this proved not to be thecase, with MLN from infected WT mice actually producing sig-nificantly more Ag-specific IL-10 than those from CD154Ϫ/Ϫ mice( p Ͻ 0.01), and with no significant difference between the groupsin the level of NO detected. Consistent with this, immunohisto-chemical staining showed that there was no elevation in iNOSlevels in lung, liver, or gut from infected CD154Ϫ/Ϫ mice com-pared with infected WT mice (not shown). Interestingly, MLNcells from infected CD154Ϫ/Ϫ mice also showed an impaired abil-ity to produce IL-4, IL-5, and IL-13, but not IL-10, IL-12, or NOin response to polyclonal stimulation with plate-bound anti-CD3mAb (not shown). Similar results were obtained from splenocytecultures stimulated with SEA or anti-CD3 (not shown).
A ROLE FOR CD154 IN Th2 INDUCTION BY S. mansoni
No significant difference in levels of production of IL-12, TNF-␣,or IFN-␥ was seen between the groups, with each of these medi-ators being produced in low amounts. Additionally, we measuredlevels of IL-10 and NO to determine whether the reduced produc-tion of IL-4, IL-5, and IL-13 might be due to enhanced levels ofthese regulatory mediators. However, this proved not to be thecase, with MLN from infected WT mice actually producing sig-nificantly more Ag-specific IL-10 than those from CD154Ϫ/Ϫ mice( p Ͻ 0.01), and with no significant difference between the groupsin the level of NO detected. Consistent with this, immunohisto-chemical staining showed that there was no elevation in iNOSlevels in lung, liver, or gut from infected CD154Ϫ/Ϫ mice com-pared with infected WT mice (not shown). Interestingly, MLNcells from infected CD154Ϫ/Ϫ mice also showed an impaired abil-ity to produce IL-4, IL-5, and IL-13, but not IL-10, IL-12, or NOin response to polyclonal stimulation with plate-bound anti-CD3mAb (not shown). Similar results were obtained from splenocytecultures stimulated with SEA or anti-CD3 (not shown).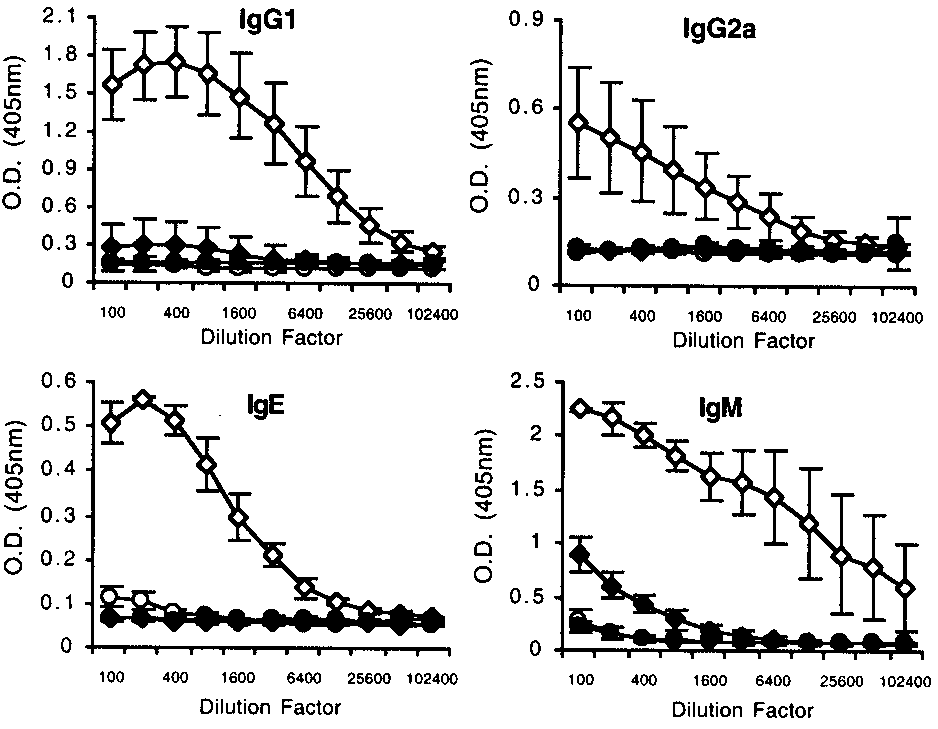 IgE, and the reduced numbers of granulocytes evident in their pe-ripheral blood and granulomas. Furthermore, this inability tomount a Th2 response has fatal consequences for infected animals.
IgE, and the reduced numbers of granulocytes evident in their pe-ripheral blood and granulomas. Furthermore, this inability tomount a Th2 response has fatal consequences for infected animals.