CHAPITRE 5 PROBLÈMES ÉVOLUTIFS La découverte de mutations capables de prolonger la vie des animaux a posé des problèmes aux évolutionnistes, qui pouvaient difficilement admettre que l’activité d’un gène puisse avoir pour effet de réduire la longévité, donc d’abaisser le potentiel reproductif des animaux. Pourtant, la liste des gènes dont les produits tendent à raccourcir la vie s’allonge sans arrêt. SOMMAIRE Page 5.1. Problèmes relatifs aux organismes unicellulaires……………………………………………………………….2
5.1.1. Protection contre la sénescence.3
5.1.2. Avantages de la sénescence………………………………………………………………………………………….3
5.2. Problèmes relatifs aux animaux…………………………………………………………………………4
5.2.1. Origine de la dichotomie soma/germen…………………………………………………………………………….4
5.2.2. Avantages de la dichotomie soma/germen……………………………………………………………………….5
5.2.3. Pérennité des systèmes anti-sénescence…………………………………………………………………………5
5.2.4. Signification de la sénescence…………………………………………………………………………………………6
5.3. Théories concernant l’origine de la sénescence………………………………………………………………….7
5.3.1. La théorie de Wal ace…………………………………………………………………………………………………….7
5.3.2. L'intuition de Haldane…………………………….……………………………………………………………………….8 .
5.3.3. Les théories modernes du vieil issement………………………………………………………………………….9
5.3.4. Évaluation des théories…………………………………………………………………………………………………10
5.3.5. Les dangers de la vie sauvage ………………………………………………………………………………………11
5.3.6. Mutations cumulatives ou pléiotropes ?.12
5.3.7. Gérontogènes instaurant la sénescence réplicative…………………………………………………………12
5.3.8. Gérontogènes instaurant la sénescence métabolique………………………………………………………13
5.3.9. Double effet des facteurs capables de prolonger la vie…………………………………………………….14
5.4. Origine de la ménopause……………………………………………………………………………………………………15
5.4.1. Théorie non adaptative…………………………………………………………………………………………………15
5.4.2. Théorie adaptative………………………………………………………………………………………………………….6
5.5. Origine de l'apoptose………………………………………………………………………………………………………….7
5.5.1. Relation entre l'apoptose et la mitose…………………………………………………………………………….17
5.5.2. Rôle des mitochondries dans l'apparition de l'apoptose.17
5.5.3. Difficultés d'interprétation…………………………………………….………………………………………………18
5.6. Conclusion……………………………………………………………………………………………………………………….19 Bibliographie………………………………………………………………………………………………………….………………19
our Darwin et la quasi-totalité des biologistes contemporains, les êtres vivants ont évolué sous
P l'aiguilon de la sélection naturele. Le critère sélectif réside dans la capacité des individus à
survivre et à engendrer des descendants nombreux, pouvant se reproduire efficacement à leur tour. Ce qui importe, c'est l'aptitude de chacun à propager ses gènes. Certains darwinistes radicaux voient dans les gènes des entités « égoïstes », ayant comme seule préoccupation de se multiplier au détriment des autres. En somme, les organismes ne seraient que des vecteurs utilisés par les gènes pour se propager. Malgré son caractère réducteur et anthropomorphique, ce point de vue reflète au moins une partie de la réalité.
L'instauration de la sénescence pose des problèmes épineux. Ce phénomène a pour effet de
restreindre la longévité des individus, donc de limiter le nombre de descendants qu'ils peuvent engendrer. Le succès reproductif étant le critère ultime de sélection, on peut se demander comment cette espèce de malthusianisme a pu apparaître et se maintenir dans diverses lignées d'êtres vivants. Cette question a suscité chez les évolutionnistes toutes sortes de réflexions visant à rendre l'instauration de la sénescence compatible avec l'orthodoxie darwinienne, aussi bien chez les organismes unicel ulaires que chez les animaux. Une particularité de l’espèce humaine a soulevé beaucoup de discussions parmi les darwinistes. Il s’agit de la ménopause, dont l’origine est difficile à comprendre, parce qu’el e abolit la fécondité des femmes bien avant le terme normal de leur vie. L'origine de l'apoptose a également fait l'objet de nombreuses spéculations.
5.1. PROBLÈMES RELATIFS AUX ORGANISMES UNICELLULAIRES On considère en général que les organismes unicel ulaires sont épargnés par les deux formes de sénescence qui menacent les cel ules des animaux. Mais ce n’est manifestement pas vrai, puisque les bactéries, la levure et les protozoaires peuvent souffrir de sénescence (chapitre 1, section 1.1). Ces constatations donnent à penser que ce phénomène existait déjà avant même que les animaux n’apparaissent. Toutefois, rien ne dit que la sénescence qui affecte les bactéries et les eucaryotes unicel ulaires a quelque chose à voir avec cel e qui condamne le soma des animaux à un dépérissement progressif. 5.1.1. PROTECTION CONTRE LA SÉNESCENCE
La plupart des bactéries ne peuvent pas souffrir de l’érosion des télomères, parce qu’el es ont des chromosomes en anneau. Les rares bactériesdont les chromosomes sont linéaires (Streptomyces, Borrelia, Rhodococcus) protègent les extrémités de ceux-ci grâce à des répétitions, non pas directes,
comme chez les eucaryotes, mais palindromiques. Mais le mécanisme d’entretien de ces répétitions n’est pas encore élucidé. Les eucaryotes unicel ulaires sont également préservés de la télotomie, car ils possèdent une télomérase qui est constamment active. La sénescence mitotique dont souffrent les cel ules mères de levure et les cel ules végétatives des ciliés* n'est pas due à des problèmes de télomères, mais à des divisions d'un type particulier, qui répartissent de manière inégale certaines macromolécules (ADN, protéines) entre les cel ules fil es.
Les bactéries semblent ignorer la sénescence métabolique. El es se protègent contre les
radicaux oxydants grâce à une superoxyde dismutase très efficace et à des polyamines, composés hydrocarbonés portant plusieurs fonctions -NH2. La sénescence métabolique n'affecte pas non plus les eucaryotes unicel ulaires. La levure ne craint pas les radicaux oxydants, puisqu'el e répond à la raréfaction du glucose en accroissant sa consommation d'oxygène (chapitre 3, section 3.6.3), ce qui n'empêche pas les cel ules mères de produire davantage de bourgeons. Un moyen de défense efficace consiste à séquestrer les protéines oxydées dans les cel ules mères, ce qui pourrait contribuer au dysfonctionnement général dont el es souffrent après une vingtaine de mitoses (section 5.1.2. AVANTAGES DE LA SÉNESCENCE
Plusieurs types d'organismes unicel ulaires compromettent la propagation de leurs gènes en limitant la capacité de prolifération de leurs cel ules ou de leurs noyaux. Cette propriété les rend moins compétitifs dans un monde qui privilégie avant tout la vitesse de propagation. El e n’aurait pas pu s’instaurer ni se maintenir si les organismes qui l’ont adoptée n’en tiraient quelque bénéfice.
La sénescence repérée chez les bactéries s’accompagne d’une asymétrie visible ou invisible
des divisions (chapitre 1, section 1.1.1). Certains auteurs pensent que la partition inégale du contenu cel ulaire est un moyen d’éliminer les macromolécules endommagées, ou trop coûteuses à remettre en état. Si l’on poursuit ce raisonnement, on peut même se demander si l’asymétrie des divisions n’est pas une conséquence plutôt qu’une cause du vieil issement inexorable de certains constituants des cel ules. Si c’était le cas, l’immortalité cel ulaire n’existerait pas vraiment.
En tout cas, l’effet protecteur des divisions asymétriques peut être invoqué chez la levure.
Apparemment, les cel ules mères préservent la viabilité et le potentiel prolifératif des bourgeons en évitant qu'ils héritent les macromolécules encombrantes ou nocives (ADN ribosomique, protéines oxydées) et les mitochondries non fonctionnel es qui s'accumulent entre les mitoses. En chargeant les cel ules mères de déchets, les divisions inégales les condamnent à mourir, mais assurent - pour un temps tout au moins - la vitalité des bourgeons.
Il est plus facile d'estimer l'avantage que procure aux ciliés, aux foraminifères et aux
phytomonadines* l'acquisition d'un soma et d'un germen, malgré la perte de l'immortalité qui en résulte pour une partie de leurs noyaux ou de leurs cel ules. En fait, la division des tâches entre soma et
germen leur a permis de construire des cel ules ou des colonies mieux armées pour survivre et se multiplier. Les ciliés et les foraminifères ont accru le volume de leur cytoplasme en confiant la gestion du métabolisme à un ou plusieurs noyaux spécialisés, qui acquièrent de nombreuses copies de chacun de leurs gènes, mais deviennent mortels. Certaines phytomonadines ont partagé leurs cel ules en deux catégories, les unes protégeant les autres pendant la période vulnérable où el es construisent de nouvel es colonies ou se transforment en gamètes. La protection que le soma offre au germen a un coût, qui est la mortalité de ses éléments.
Puisqu'el e revêt des modalités très variées, il semble que la dichotomie soma/germen et la
mortalité qu'el e entraîne sont apparues à plusieurs reprises dans diverses lignées d'eucaryotes : au moins trois fois chez les unicel ulaires (ciliés, foraminifères, phytomonadines), et une fois encore chez les animaux.
5.2. PROBLÈMES RELATIFS AUX ANIMAUX
L'origine des animaux reste mystérieuse. Beaucoup de chercheurs pensent qu'ils se sont formés il y a plus d'un mil iard d'années à partir de protozoaires flagel és. Ils fondent leur conviction sur l'existence d'un certain nombre de caractères communs entre les représentants actuels de ces deux groupes. En fait, les animaux ont reçu de leurs ancêtres unicel ulaires un vaste héritage, dont on retrouve des traces dans les lignées col atérales issues des eucaryotes les plus anciens.
L'inventaire de l'héritage n'est pas facile à établir. Il faut évidemment y inclure tous les gènes qui
gouvernent l'organisation des chromosomes, l'exécution et le contrôle de la mitose et de la méiose, le fonctionnement et la multiplication des mitochondries, etc. D'autres caractères semblent exclus de l'héritage parce qu'ils sont propres aux animaux. Apparemment, ceux-ci ont inventé la dichotomie soma/germen et les mécanismes vouant leur soma à la mortalité. Toutefois, les choses ne sont pas aussi simples, parce que certains processus garantissant l'immortalité des eucaryotes unicel ulaires ont été conservés par les animaux, mais remaniés de tel e façon qu'ils déterminent chez eux la longévité du soma, comme nous al ons le voir (section 5.2.3).
5.2.1. ORIGINE DE LA DICHOTOMIE SOMA/GERMEN
Puisqu'el e existe chez presque tous les animaux, il est probable que la dichotomie soma/germen s'est instaurée dès le début de leur évolution et maintenue dans la plupart des groupes. Mais il n'est pas exclu qu'el e existait déjà chez les protozoaires fondateurs, sous la forme de deux catégories de noyaux, les uns voués à la gestion des affaires courantes, les autres à la propagation des gènes, comme cela se passe notamment chez les ciliés (annexe II).
La formation du soma correspond à la phase de multiplication végétative des protozoaires, à
cela près que les cel ules produites par l'œuf quand il se segmente ne se dispersent pas, mais restent associées et construisent un embryon. La formation du germen réunit les deux types de reproduction (végétative et sexuée) qui alternent au cours du cycle vital des protozoaires. El e commence au cours du développement par une série de mitoses et se termine chez l'adulte par la différenciation des gamètes. De la sorte, le germen prolonge à travers chaque individu la lignée d'êtres vivants qui se sont succédé sans interruption, tout en évoluant sans cesse, depuis près de quatre mil iards d'années.
5.2.2. AVANTAGES DE LA DICHOTOMIE SOMA/GERMEN
L'acquisition d'un soma « jetable » a favorisé l'apparition d'animaux de plus en plus complexes. Débarrassées de la fonction reproductrice, les cel ules somatiques ont pu s'engager de manière irréversible dans un processus de différenciation, créateur de fonctions nouvel es, parce qu'el es ne devaient plus se convertir en gamètes. Certaines cel ules renoncent définitivement à se diviser quand el es se différencient, parce que leur état leur interdit de réaliser les transformations morphologiques nécessaires à l'accomplissement de la mitose. On peut dire que la vie n'a pu évoluer vers les formes complexes que nous connaissons à présent, sans inventer la mort.
Parmi les fonctions prises en charge par le soma des animaux, la nutrition revêt une importance
primordiale, parce c'est pour eux le seul moyen de se procurer les molécules organiques indispensables à la construction de leurs cel ules, ainsi qu'à la production d'énergie. Pour se nourrir, les animaux ont organisé leur soma autour d'une cavité ouverte sur l'extérieur, où ils introduisent et digèrent les proies qu'ils peuvent capturer. Ce mode de vie crée un danger permanent, car tout prédateur peut devenir à son tour une proie pour un organisme plus grand, plus agile ou mieux armé que lui.
La prédation a instauré une lutte pour la vie, qui constitue un des moteurs de l'évolution. Seules
ont prospéré les lignées d'individus qui ont réussi à se reproduire, donc à transmettre leurs gènes, avant de succomber à la voracité de leurs ennemis naturels, ou à tout autre péril : infection bactérienne ou virale, disette prolongée, sécheresse, chaleur ou froid excessif.
5.2.3. PÉRENNITÉ DES SYSTÈMES ANTI-SÉNESCENCE
Les eucaryotes les plus divers utilisent deux systèmes généraux de protection contre la sénescence. On peut donc penser que ces systèmes ont été mis au point par un représentant très ancien de ce groupe et conservés par les innombrables générations qui ont suivi. La plupart des animaux continuent à les employer, mais certains y ont introduit des modifications. Ils se protègent contre la sénescence réplicative par l’action de la télomérase. Ils se protègent contre la sénescence métabolique grâce à la sirtuine. Le gène de la télomérase confère l'immortalité ; celui de la sirtuine a le pouvoir de prolonger la vie.
En adoptant pour leurs chromosomes une structure linéaire, les eucaryotes ont condamné leurs
cel ules à souffrir de la sénescence réplicative, car les télomères ont une fâcheuse tendance à fusionner et à raccourcir. Pour combattre ces fléaux, ils ont adapté un système préexistant, qui servait à réparer les cassures db dans l’ADN. Le dimère Ku**, les kinases DNA-PK** et ATM**, le complexe MRN** et la protéine TRF2 témoignent de ce bricolage évolutif, car tous participent à des degrés divers, non seulement à la réparation de l’ADN (annexe VII.B), mais aussi l’entretien des télomères. Pour compenser l’érosion de leurs chromosomes, les eucaryotes ont réutilisé la rétrotranscriptase*, un enzyme très ancien, dont l’origine remonte probablement à l’époque où l’ADN a supplanté l’ARN en tant que vecteur de l’information génétique. Tous les groupes d'eucaryotes (protozoaires, champignons, plantes vertes, animaux) utilisent la télomérase pour se protéger contre la télotomie. Partout, l’enzyme comporte deux sous-unités de nature différente (un ARN et une protéine), et ajoute
des séries de nucléotides à l’extrémité de l’ADN, ce qui crée des queues sb, riches en guanylates. Toutefois, certains animaux ont abandonné ce système et en ont adopté un autre, qui présente malgré tout des similitudes avec le système ancestral. Les insectes diptères, et en particulier la drosophile, recourent à des rétrotransposons* pour maintenir l'intégrité de leurs chromosomes. Ces éléments mobiles sautent de télomère en télomère et propagent des séquences répétées. La transposition fait intervenir un ARN et une rétrotranscriptase, comme le système d'entretien des télomères employé par la majorité des eucaryotes.
La levure, C. elegans, la drosophile et les mammifères utilisent la sirtuine pour retarder ou
ralentir la sénescence de leurs cel ules. Cette protéine agit partout sur les histones, mais el e a des fonctions assez différentes chez ces divers organismes (chapitre 4, section 4.2.2). La protéine Sir2 de la levure semble fonctionner uniquement comme répresseur de transcription. La sirtuine des mammifères (Sirt1) se comporte, non seulement comme répresseur, mais aussi comme activateur de transcription. Apparemment, el e s’est procuré une nouvel e cible au cours de l’évolution de ces animaux (la protéine P53), ce qui lui permet d’agir à la fois sur la sénescence des cel ules et sur l’apoptose.5.2.4. SIGNIFICATION DE LA SÉNESCENCE
Les animaux réservent à leur soma les inconvénients de la sénescence. Cel e-ci peut s'interpréter comme une restriction imposée par la vie en société, qui empêche les cel ules de se diviser trop de fois ou de vivre trop longtemps.
Pour les cel ules somatiques, la sénescence n'est pas une fatalité, mais l'effet d'un acte
« volontaire », qu'el es accomplissent dès le début de leur vie. Cet acte requiert la mise en activité de plusieurs gènes, dont quelques-uns ont été identifiés. C’est ce qui explique le caractère dominant de la mortalité sur l’immortalité, révélée par les expériences de génétique somatique (chapitre 2,sections 2.5.10 et 2.5.17). En ce qui concerne la sénescence réplicative, les produits des gènes conférant la
mortalité ont pour cible le gène de la télomérase (tert), qu'ils répriment (chapitre 2, section 2.5.12). Si l'un de ces gènes est muté, les cel ules somatiques retournent à l'état d'immortalité propre aux organismes unicel ulaires. Les gènes qui instaurent la sénescence métabolique réduisent la résistance des cel ules aux dérivés nocifs de l'oxygène. Si l'un d'entre eux est muté, la résistance se renforce et les animaux vivent plus longtemps (chapitre 3, section 3.10). Notons encore que les cel ules en division sont naturel ement sensibles aux radicaux oxydants, parce qu’el es ralentissent, par l’intermédiaire du facteur FoxO3**, la synthèse de plusieurs enzymes de désintoxication
Ces remarques conduisent à une conclusion qui peut sembler paradoxale : dès le début de leur
vie, les animaux préparent activement leur fin en limitant la capacité de prolifération de leurs cel ules, ainsi que leur aptitude à limiter les dégâts provoqués par les radicaux oxydants. Les gérontogènes responsables de ces effets agissent en permanence et leur activité n'augmente pas nécessairement avec l'âge. C'est ce qu'a montré une étude par la technique des biopuces sur l'ensemble des gènes de
C. elegans : les produits des gènes daf2/inr** et age1/daf23* figurent parmi ceux que l'on trouve en
quantités constantes dans les cel ules des animaux jeunes ou âgés.
5.3. THÉORIES CONCERNANT L’ORIGINE DE LA SÉNESCENCE
La découverte des gérontogènes a plongé certains biologistes dans la perplexité. Mais les évolutionnistes se sont vite ressaisis. À vrai dire, ils n'avaient pas attendu les années 1980 pour réfléchir sur l'origine de la mortalité et construire des théories visant à expliquer ce phénomène à la lumière de la sélection naturel e. La première de ces théories a vu le jour avant même que les conceptions de Darwin ne connaissent le succès. 5.3.1. LA THÉORIE DE WALLACE
Concernant l'origine de la sénescence, Alfred Russel Wal ace, co-inventeur avec Darwin de la théorie de la sélection naturel e, fit une proposition qui peut sembler pertinente, reprise quelques années plus tard par August Weismann, le fondateur de la théorie du plasme germinal. Selon ces auteurs, la mort des individus favorise le succès de l'espèce dont ils font partie. La limitation de la longévité accélère le renouvel ement des générations, ce qui facilite l'adaptation des animaux à des modifications rapides de l'environnement. En se condamnant à mourir, les individus âgés aideraient les jeunes à survivre, parce qu'ils n'entrent pas en compétition avec eux pour l'espace et les ressources alimentaires, qui sont toujours limités. Mais cette explication teintée d'idéologie ne résiste pas à un examen sérieux. On peut lui opposer au moins deux arguments.
Le premier argument relève de l'observation. Il ne semble pas que la sénescence limite tant soit
peu la durée de vie des animaux sauvages. La plupart meurent prématurément d'accidents divers. On estime que les souris de nos régions atteignent rarement l'âge de deux ans. Plus de 90 % succombent dans leur première année, avant d'éprouver les premières atteintes de la vieil esse.
Le second argument est de nature théorique. L'idée de Wal ace et de Weismann fait appel aux
notions d'altruisme* et de sélection de groupe*. Les théories faisant appel à ces notions soulèvent des difficultés parce qu’el es sont confrontées au problème des « tricheurs ». Dans le cas particulier du vieil issement, si des mutations conféraient aux tricheurs l’aptitude à vivre plus longtemps, ils pourraient tirer parti du sacrifice des autres membres de leur groupe. Mais à supposer que de tel es mutations se répandent dans les populations animales, el es devraient tôt ou tard supprimer l’avantage dont les tricheurs bénéficiaient au début. De toute façon, les darwinistes soutiennent que la sélection naturel e ne travail e pas pour le bien des espèces, mais pour celui des individus, ou plutôt de leurs descendants. El e tend à accroître la fréquence des gènes propagés par les individus les plus aptes, au détriment des gènes propagés par les individus les moins adaptés. Si un animal limite la durée de sa vie, il contribuera peut-être à propager ses gènes, en faisant « place nette » pour sa progéniture. Mais on peut difficilement imaginer comment il pourrait faire en sorte que sa mort ne profite qu'à ses descendants, plutôt qu’à ses concurrents, apparentés ou non, honnêtes, ou tricheurs.
Seuls quelques rares animaux réservent à leurs descendants le bénéfice de leur mort. Ils se
laissent tout simplement dévorer par eux. Ce phénomène, appelé matriphagie, est assez rare, mais Aristote le connaissait déjà. Il se rencontre chez certains arachnides, animaux réputés pour leur propension au cannibalisme, pratiqué par les femel es de plusieurs espèces au détriment des mâles.
Chez les araignées sociales appartenant au genre Diaea, les jeunes sucent l'hémolymphe de leur mère au niveau des articulations de ses pattes, puis la dévorent entièrement. Chez les acariens des genres Siteroptes, Acarophenax et Adactylidium, les femel es ne pondent pas. Les embryons éclosent dans leur corps : les larves croissent en mangeant les organes internes de leur mère, qui finit par périr, laissant subsister une carcasse vide, que sa progéniture, devenue adulte, s'empresse de quitter. Mais ces cas extrêmes d'altruisme maternel ne suffisent pas pour rendre crédible l'idée de Wal ace et Weismann.
5.3.2. L'INTUITION DE HALDANE
Pour de nombreux évolutionnistes contemporains, la durée de la vie ne constitue pas un caractère adaptatif, de nature à influencer la propagation des gènes dans un environnement déterminé. El e n'est ni avantageuse, ni désavantageuse, mais neutre, parce qu'el e n'est que la conséquence d'une sélection opérant sur d'autres caractères. Tout le problème est de savoir quels sont ces caractères.
En 1941, John Haldane fit une remarque judicieuse à propos de la sénescence qui compromet
tôt ou tard la survie des êtres humains et des animaux. Sa réflexion se fondait sur les propriétés d'une maladie héréditaire qui entraîne chez l'homme une dégénérescence progressive du cerveau. Il s'agit de la chorée de Huntington*. Cette maladie présente deux caractères exceptionnels. En premier lieu, el e est due à une mutation dominante. En second lieu, el e frappe ses victimes à un âge assez avancé (35 à 45 ans), lorsque la plupart ont déjà eu des enfants, dont la moitié a reçu le gène altéré et développera la maladie. D'habitude, les mutations dominantes à effet délétère disparaissent rapidement des populations humaines ou animales, car el es réduisent l’aptitude des individus à se reproduire. Mais ce n'est pas le cas en ce qui concerne la chorée de Huntington, qui affecte environ un nouveau-né sur 10 000 en Europe, en Amérique et en Australasie. Manifestement, l'évolution ne peut pas purger les populations des gènes létaux si les effets des mutations qu'el es ont subies se manifestent après la période de reproduction.
Partant de cette idée, Haldane se demanda comment l'évolution peut agir sur les mutations qui
exercent leurs effets encore plus tardivement que la chorée de Huntington. D'après lui, l'évolution peut difficilement éliminer de tel es mutations. Prenons l'exemple de la drosophile. Toute mutation qui diminuerait la vitalité des mouches après l'âge de trois semaines n'a aucune chance d'être extirpée des populations naturel es (chapitre 4, section 4.6.1). En fait, la sélection agit sur les individus avec une force décroissante à mesure qu'ils vieil issent, parce que le nombre de descendants qu'ils peuvent engendrer diminue constamment. Devenu stérile, un individu perd toute valeur du point de vue évolutif. Sa disparition sera sans conséquence pour l'avenir de sa lignée - à moins qu'il ne reste quelque temps encore utile à la survie de sa progéniture.5.3.3. LES THÉORIES MODERNES DU VIEILLISSEMENT
Trois théories s'inspirent plus ou moins directement des conceptions de Haldane. El es essaient d'expliquer comment la sélection naturel e a pu retenir des mutations qui ont modelé des gènes capables d'écourter la vie des animaux, ce qui paraît impossible. Leurs formulations se ressemblent, car el es font appel à des mutations agissant de manière variable suivant l'âge des individus
(tabl. 5.1).
La théorie du soma jetable invoque un compromis que les animaux doivent maintenir entre
deux exigences fondamentales : l'entretien du corps et la reproduction. Les animaux consacrent l'énergie dont ils disposent à maintenir leur soma en bonne condition, mais pas au-delà d'une limite fixée par leurs chances de survie dans la nature. Passé cette limite, l'entretien du soma devient inutile. Il peut sans inconvénient disparaître. En somme, la sélection naturel e agit sur les animaux en favorisant leur survie jusqu'à ce que la reproduction soit assurée. Une fois cette tâche accomplie, le sort du soma ne l'intéresse plus. Cela revient à dire que, du point de vue sélectif, la prolongation de la vie au-delà d'une limite fixée par le milieu naturel n'offre aucun avantage. Chez certains animaux semelpares, comme les saumons, le soma dépérit sitôt la reproduction assurée.
Les deux autres théories du vieil issement font appel à des mutations ayant pour effet de
promouvoir la sénescence après un certain âge, ce qui est la marque distinctive des gérontogènes. Toutes deux supposent que la sélection naturel e accroît la fréquence de certaines mutations qui apparaissent spontanément chez les animaux, bien qu'el es aient pour conséquence d'abréger leur vie. Ces mutations ont été retenues parce qu’el es accroissent d’une manière ou d’une autre le succès reproductif des animaux qui les ont subies.
L'une de ces théories invoque des mutations purement délétères, qui s'accumulent dans les
populations animales et additionnent leurs effets (fig. 5.1). La sélection se contenterait d'éliminer plus efficacement les mutations qui compromettent la viabilité des individus jeunes que cel es qui affaiblissent les individus âgés. La dernière théorie fait appel à des mutations pléiotropes*, ayant un effet bénéfique au début de la vie, mais préjudiciable à la fin. La sélection conserverait les mutations
diminuant la vitalité les individus devenus vieux, pourvu qu'el es augmentent les chances de survie des jeunes. Suivant ce point de vue, la longévité des animaux serait fixée par un subtil équilibre entre les effets contradictoires d'un certain nombre de gérontogènes.
Fig. 5.1. Modes d'action des gérontogènes. A. Suivant la théorie des mutations cumulatives, les gérontogènes n'agissent que sur les individus âgés. B. Suivant la théorie des mutations pléiotropes, les gérontogènes agissent à la fois sur les jeunes et sur les
vieux, renforçant la vitalité des uns, mais affaiblissant cel e des autres. Comme les animaux se reproduisent
avec une efficacité décroissante au cours de leur vie, tout effet défavorable se manifestant après la période de
fécondité peut difficilement être éliminé par la sélection naturel e, parce que les mutations produisant cet effet
ne sont pas transmissibles. Par ce mécanisme, les populations d’animaux animales auraient intégré un certain
nombre de gérontogènes qui fixent la durée de leur vie.
5.3.4. ÉVALUATION DES THÉORIES
Les trois théories expliquent de façon satisfaisante la manière dont les gènes contrôlent le début et la fin de la vie. Les chromosomes contiennent un « programme » de développement très détail é, sous la forme d'un ensemble de gènes qui gouvernent de manière rigoureuse le modelage de l'embryon et le fonctionnement de ses cel ules. Mais ils ne renferment aucun « programme » de mort qui ferait disparaître l'organisme à un moment précis et d'une manière prévisible. L'œuf « sait » par avance comment bâtir un organisme apte à propager efficacement ses gènes, mais ne « sait » pas comment le déconstruire de manière ordonnée, une fois cette tâche accomplie. Tout cela se comprend, parce que la sélection naturel e n'intervient plus après la période de reproduction. El e a donc été incapable de retenir les mutations qui auraient permis d'élaborer un programme de mort qui fonctionnerait de la même façon chez tous les animaux d’un même groupe.
Il reste à comparer les trois théories du vieil issement. Comme el es se réfèrent à des
événements survenus dans un passé lointain, il est très difficile de réaliser des expériences qui pourraient les confirmer ou les infirmer. Faute de mieux, essayons de voir comment les théories s'accordent avec les observations glanées dans la nature et avec les propriétés des gérontogènes que l'étude des animaux de laboratoire a permis de découvrir (chapitre 3, section 3.10).
5.3.5. LES DANGERS DE LA VIE SAUVAGE
La théorie du soma jetable s'inspire des observations, déjà mentionnées, concernant la longévité des animaux sauvages. Revenons aux souris naines pit1** ou prop1** (chapitre 3, section 3.9.1). Imaginons qu'une mutation retarde la sénescence d'un individu sauvage, en réduisant la sécrétion d'hormones par l'hypophyse ou le foie. Une tel e mutation n'a guère de chances d'être adoptée par l'ensemble des souris, car les mutantes seront mortes - le plus souvent de froid, étant donné leur tail e réduite - bien avant d'avoir pu profiter de la prolongation de la vie dont el es pourraient jouir.
La théorie a été utilisée pour essayer de comprendre pourquoi les animaux qui sont exposés
durant toute leur vie à de grands périls ont une longévité potentiel e inférieure à cel e des animaux qui en sont protégés grâce à leur tail e, leur agilité ou leur intel igence. Suivant ce point de vue, les souris vieil issent rapidement parce que la sélection n’a pas éliminé chez leurs ancêtres les mutations ayant pour effet de réduire l'efficacité des systèmes de protection contre la sénescence, comme celui qui mobilise la Parp** (section 3.3.4). Dès le début de leur vie, les dégâts causés par les radicaux oxydants s'accumulent et réduisent peu à peu la viabilité du soma. On peut penser que les oiseaux et les chéiroptères vivent longtemps parce que le vol les protège mieux contre les prédateurs que les vertébrés terrestres, et qu'ils ont conservé ou développé des systèmes anti-sénescence efficaces.
Cette proposition semble raisonnable. Cependant, el e est contredite par des mesures faites sur
des poissons de rivière (guppys) qui vivent dans un environnement plus ou moins dangereux, c’est-à-dire plus ou moins riche en prédateurs. Les animaux les plus exposés à la prédation, qui meurent donc en moyenne plus jeunes, ont en aquarium une longévité supérieure à cel es des animaux les moins exposés. Les auteurs de l’étude appel ent à une révision de la théorie. Le moins qu’on puisse dire est qu’el e ne paraît pas valable pour la totalité des animaux.
Quoi qu’il en soit, la notion incluse dans la théorie du soma jetable offre un moyen d'évaluer
correctement le succès reproductif des animaux. Reprenons l'exemple des mutants daf2/inr** de
C. elegans. Les hermaphrodites normalement constitués pondent près de 300 œufs au cours de leur
vie, tandis que les mutants daf2 en pondent 200 à 250, mais en 40 jours, au lieu de 20. Si les animaux à vie prolongée étaient relâchés dans la nature, leur succès reproductif serait moindre que celui de leurs congénères normaux, car ils risqueraient davantage de périr avant d'avoir donné naissance à tous les descendants qu'ils peuvent engendrer. En limitant la durée de vie des animaux, sans accroître notablement leur fécondité, le gérontogène daf2 a donc pour effet paradoxal d'augmenter leur potentiel reproductif.
5.3.6. MUTATIONS CUMULATIVES OU PLÉIOTROPES ?
Les deux dernières théories ont inspiré de nombreuses expériences, menées principalement sur la drosophile. Les résultats sont en total désaccord. Ils soutiennent l'une ou l'autre théorie, suivant la nature des facteurs étudiés pour l'influence qu'ils exercent sur la longévité des mouches : fécondité, croisements consanguins, etc. Leur interprétation fait appel aux subtilités de la génétique des populations, peu accessibles aux non-spécialistes.
À défaut de départager les deux camps sur des bases expérimentales, essayons de voir s'il
existe des gérontogènes répondant aux spécifications de l'une ou de l'autre théorie. Si un gérontogène obéit aux critères de la théorie des mutations cumulatives, son inactivation devrait prolonger la vie, sans affecter les individus avant leur période de pleine maturité sexuel e (section 5.3.3, fig. 5.1). S'il se conforme aux critères de la théorie des mutations pléiotropes, un renforcement de son activité devrait avoir des avantages pour les jeunes, mais des inconvénients pour les vieux. Inversement, une mutation prolongeant la vie devrait avoir certains désavantages pour les jeunes.
En fait, beaucoup de gérontogènes se comportent donc comme le prévoit la théorie des
mutations pléiotropes. On peut le vérifier en examinant, comme nous al ons le faire, les propriétés des gènes qui favorisent l'une et l'autre forme de sénescence. Par ail eurs, les traitements qui prolongent la vie des animaux ont aussi des inconvénients pour ceux qui les subissent. Ici encore, une certaine forme de pléiotropie apparaît. Mais ces observations ne permettent pas de rejeter en bloc la théorie des mutations cumulatives. De fait, une de ses principales implications - l'effet additif de nombreux gènes sur la détermination de la durée de la vie - a été vérifiée chez C. elegans par l'analyse détail ée de l'activité des gènes contrôlés par le produit de daf2/inr.
5.3.7. GÉRONTOGÈNES INSTAURANT LA SÉNESCENCE RÉPLICATIVE
Chez l’homme et la souris, la protéine P53 protège contre le cancer, mais tend à accélérer la sénescence. C’est ce qu’a révélé l’étude d’un polymorphisme au niveau du gène P53 humain : les personnes portant deux al èles « forts » du gène (génotype P5372R/R) souffrent moins fréquemment de cancer que cel es qui portent deux al èles « faibles » (génotype P5372P/P), mais meurent en moyenne plus tôt (chapitre 2, section 2.10.3). De même, les souris dont la protéine P53 a une activité renforcée (génotype p53+/m) souffrent moins souvent de cancer que les témoins, mais vieil issent plus vite. Inversement, les animaux de génotype p53-/- devraient vivre plus longtemps, s’ils pouvaient échapper aux nombreux cancers que l’absence de protéineP53 laisse rapidement se développer.
Dans les deux cas évoqués ci-dessus, le gène p53 agit conformément à ce que prévoit la
théorie de la pléiotropie antagoniste. Son action a un effet favorable pour les individus jeunes, puisqu’el e les protège contre le cancer, mais défavorable pour les vieux, puisqu’el e les fait mourir plus rapidement. En somme, la protéine P53, ainsi que d’autres mécanismes limitant la prolifération cel ulaire, empêcheraient les animaux (et les êtres humains) de mourir jeunes d’un excès de cel ules fonctionnant mal, mais les ferait peu à peu dépérir d’un déficit de cel ules fonctionnant bien. Si ces mécanismes sont inopérants, des cancers peuvent apparaître dès le plus jeune âge, comme cela se produit chez les enfants qui portent une mutation dans le gène RB. Plus tard dans la vie, les fonctions
de l’organisme se détériorent, parce que le renouvel ement de certaines cel ules souches* n’est plus assuré. La sénescence réplicative imposée aux cel ules somatiques serait donc essentiel ement un mécanisme suppresseur de tumeur.
Mais cette proposition soulève des difficultés. Continuons à raisonner au sujet de la protéine
P53. Toute augmentation de son activité ou de sa concentration intracel ulaire n’entraîne pas nécessairement une accélération de la sénescence (chapitre 2, section 2.10.3). Par ail eurs, la protéine P53 n’agit pas chez tous les animaux comme suppresseur de tumeur. Chez C. elegans et la drosophile, son rôle se limite au déclenchement de l’apoptose. Au surplus, le risque pour ces animaux de développer un cancer est très restreint, voire inexistant, parce que leur vie est brève et que leur soma se compose en totalité ou en majorité de cel ules qui ne se divisent plus.
5.3.8. GÉRONTOGÉNES INSTAURANT LA SÉNESCENCE MÉTABOLIQUE
Parmi les mutations susceptibles de ralentir la sénescence métabolique, beaucoup ont des effets défavorables, qui se manifestent dès la métamorphose ou la naissance, et même plus tôt. Les mutants ont une croissance ralentie, souffrent d’une diminution de la fécondité, de torpeur, d’une carence alimentaire ou de nanisme (tabl. 5.2). Du point de vue sélectif, les mutants sont donc moins aptes à survivre que les animaux de génotype normal. Relâchés dans la nature, ils devraient rapidement disparaître, sans laisser de descendants. Une expérience réalisée en laboratoire sur C. elegans étaie ce point de vue. Si des mutants daf2 à vie prolongée sont élevés en présence
d’animaux sauvages, ils disparaissent en quatre générations, même si l’apport en nourriture est suffisant.
Puisque les mutations qui ralentissent la sénescence ont des inconvénients, c’est que les
gérontogènes qu’el es neutralisent ont un effet favorable au début de la vie. L’avantage que procure l’activité de ces gènes consiste à stimuler la croissance des jeunes. Parmi les gérontogènes découverts par mutagenèse aléatoire ou dirigée, près de la moitié agit en facilitant la pénétration du glucose dans les cel ules par l’intermédiaire de la voie de transduction* contrôlée par l’insuline ou les IGF : daf28, daf2/inr, age1/daf23*, pdk1** et sgk**1 chez C.elegans ; inr et chico/irs chez la drosophile ; pit1**, prop1**, ir**, ghr**, igfr1** et shc1** chez la souris (chapitre 3, section 3.10). Mais l’activité de ces gènes diminue la résistance des cel ules aux dégâts provoqués par les radicaux oxydants, ce qui accélère le vieil issement.
5.3.9. DOUBLE EFFET DES FACTEURS APTES À PROLONGER LA VIE
Deux types de traitements ont sur la longévité des animaux des effets contrastés, comme les mutations pléiotropes qui ralentissent la sénescence. Il s’agit de la restriction calorique et la stérilisation. Le premier type de traitement est efficace chez de nombreux animaux. Il prolonge la vie et peut, de ce fait, accroître l’efficacité de la reproduction, mais retarde la croissance. La stérilisation n’a été expérimentée avec succès que chez C. elegans et la drosophile. El e permet aux animaux de vivre plus vieux, mais au prix de la perte de la fonction essentiel e du point de vue évolutif.
Comme beaucoup de mutations qui prolongent la vie, la privation de nourriture ferme
partiel ement la voie de transduction commandée par le récepteur INR. Cette coïncidence n’est pas le fruit du hasard. En fait, la carence alimentaire gouverne un mécanisme crucial, qui optimise la reproduction en dépit des fluctuations que subit l’apport en éléments nutritifs. Ce système fonctionne chez de nombreux organismes hétérotrophes*.
La levure a adopté un tel système, puisque la privation de glucose oblige les cel ules mères à
grossir moins vite et à se diviser moins souvent, mais leur permet de le faire davantage de fois
(chapitre 2, section 3.4.1). La restriction calorique a un autre effet favorable : el e prolonge aussi la
survie des cel ules en phase stationnaire, qui peuvent attendre plus longtemps que l’abondance revienne pour recommencer à bourgeonner. Les cel ules aptes à mettre en œuvre de tel es compensations ont une descendance plus nombreuse que les cel ules incapables de le faire, ce qui constitue un avantage décisif. Il convient de rappeler que la restriction calorique agit par l’intermédiaire d’une protéine (la sirtuine) qui a des effets opposés sur les deux types de sénescence (chapitre 3, sections 3.6.5 et 3.6.6). El e retarde la sénescence réplicative, parce qu’el e augmente la capacité de
reproduction des cel ules durant la période de croissance exponentiel e, mais accélère la sénescence chronologique, en réduisant leur capacité de bourgeonner de nouveau durant la phase stationnaire.
Les animaux soumis à une carence alimentaire modérée se reproduisent plus tardivement,
mais vivent plus vieux. Si l’abondance revient, ils pourront engendrer une progéniture plus nombreuse que leurs congénères incapables d’augmenter la durée de leur vie quand la nourriture devient rare. Cela montre que les animaux ne subissent pas la sénescence métabolique comme une fatalité. Ils ont acquis le moyen de la retarder lorsque les ressources alimentaires deviennent moins abondantes, ce qui préserve leur potentiel reproductif. Certains d’entre eux, tels que C. elegans, ont perfectionné cette stratégie en arrêtant les progrès de la sénescence tant que les ressources alimentaires ne leur permettent pas de croître normalement (chapitre 3, section 3.4.2).
Chez C. elegans, la perte du pouvoir reproducteur due à l’élimination du germen prolonge la vie
des adultes grâce à un système de communication entre cel ules faisant appel à une hormone hydrophobe (section 3.7.9). L’avantage d’un tel mécanisme paraît évident. Si l’appareil génital tarde à devenir fonctionnel, les animaux tendent à vivre plus longtemps, ce qui accroît leurs chances de devenir féconds.
5.4. ORIGINE DE LA MÉNOPAUSE Dans toutes les communautés humaines, les femmes deviennent stériles vers l’âge de 50 ans. Même dans les tribus africaines de chasseurs-cueil eurs, censées perpétuer un mode de vie primitif, de nombreuses femmes dépassent l’âge de 60 ans et participent activement à la col ecte de nourriture. La prolongation de la vie au-delà de la période de fécondité n’est donc pas propre aux sociétés dites industriel es.
Les évolutionnistes ont des opinions divergentes concernant l’origine de la ménopause. Les uns
dénient à ce phénomène toute valeur adaptative. Les autres estiment au contraire que son apparition a renforcé le succès reproductif de la lignée de primates dont dérivent les êtres humains actuels.
5.4.1. THÉORIE NON ADAPTATIVE
Les partisans de cette théorie considèrent la ménopause comme une conséquence inéluctable du vieil issement. Il s’agirait d’un processus que la sélection naturel e a laissé apparaître car el e n’agit pas sur les mutations qui compromettent le fonctionnement de l’organisme après un certain âge. L’apparition de la ménopause serait liée à l’extrême dépendance des jeunes enfants vis-à-vis de leurs parents, et en particulier de leur mère, pour se procurer les produits de la chasse ou de la cueil ette. En matière de nutrition, l’autonomie ne devient effective qu’après l’âge de 10 ans. Les femmes doivent donc s’occuper pendant plusieurs années de leurs enfants pour assurer leur survie. Comme la grossesse et l’accouchement entraînent des dangers croissant à mesure que les femmes vieil issent, la prolongation de la fécondité au-delà d’une certaine limite augmente pour chacune d’el es le risque de laisser des orphelins incapables de subvenir à leurs besoins alimentaires. Bien qu’el e semble avoir pour effet de diminuer le nombre total d’enfants qu’une femme peut engendrer au cours de sa vie, la ménopause n’a pas en fin de compte aucun impact sur la fécondité globale. El e serait donc neutre du point de vue sélectif.
Il est évidemment impossible de faire des expériences pour évaluer le bien-fondé de cette
théorie. Certains biologistes font des simulations reposant sur des données démographiques censées caractériser les sociétés primitives de chasseurs-cueil eurs. Mais la valeur de ces spéculations est difficile à estimer. Par exemple, il suffit pour ruiner le raisonnement esquissé ci-dessus d’admettre que les orphelins étaient pris en charge par leurs proches parents, ce qui leur évitait de mourir avant de pouvoir assurer leur subsistance.
D’autres biologistes se tournent vers la nature et comparent la manière de se reproduire des
mammifères sauvages avec cel e des êtres humains. Un groupe de chercheurs a observé pendant près de 30 ans les lions et les babouins vivant dans un parc naturel de Tanzanie. Il constate que les femel es de ces animaux éprouvent une diminution abrupte de leur fécondité, vers l’âge de 13 ans pour les unes et de 20 pour les autres. Après avoir pris soin de leur dernier jeune pendant plus d’un an (chez les lions) ou de deux (chez les babouins), el es prennent leur « retraite » et vivent encore plusieurs années. Les chercheurs notent aussi que les lionceaux ou les jeunes babouins qui ont perdu leur mère pendant leur première année ont des chances de survie très réduites. Ils concluent que la ménopause n’est pas une caractéristique propre aux femmes. À leurs yeux, la dépendance des jeunes vis-à-vis de leur mère suffit pour expliquer pourquoi les femel es vivent en général assez longtemps après avoir cessé de se reproduire. Mais d’autres auteurs, observant des populations naturel es de chimpanzés, nient l’existence d’une ménopause chez ces animaux, parce que le taux de fécondité des femel es diminue paral èlement à leur chances se survie et qu’el es continuent à se reproduire jusqu’à un âge avancé.
5.4.2. THÉORIE ADAPTATIVE
Pour les tenants de cette théorie, la ménopause aurait augmenté le succès reproductif des femmes parce qu’el e leur laisse le loisir, non seulement d’élever leurs propres enfants, mais aussi de s’occuper de leurs petits-enfants, ce qui accroît les chances de survie de ceux-ci, tout en déchargeant leurs fil es des soins que réclament les enfants en bas âge.
De nouveau, il est difficile d’évaluer la pertinence de ce raisonnement, parce qu’on ne dispose
pas de données fiables concernant les sociétés primitives de chasseurs-cueil eurs. Faute de mieux, une double équipe de démographes a essayé de mesurer l’importance de l’« effet grand-mère » dans
deux populations très différentes, l’une vivant en Finlande aux XVIIIe et XIXe siècles, l’autre au
Québec vers la fin du XIXe siècle. Il s’agissait de dénombrer les petits-enfants qu’ont eus les femmes ayant dépassé la cinquantaine, en fonction de l’âge où el es sont mortes. Il apparaît que les femmes qui ont vécu le plus longtemps ont eu la descendance la plus nombreuse. Par exemple, les finlandaises qui sont mortes vers 50 ans ont eu en moyenne 10 petits-enfants, mais cel es qui ont atteint l’âge de 80 en ont eu cinq de plus. Cette différence pourrait simplement signifier que les femmes qui ont atteint l’âge le plus avancé ont eu des fils ou des fil es robustes, aptes à engendrer de nombreux descendants. Mais cette explication n’est pas entièrement valable. En effet, les auteurs observent que les grands-mères qui vivaient dans le même vil age que leurs fils ou leurs fil es ont eu davantage de petits-enfants que cel es qui vivaient loin d’eux, c’est-à-dire à 20 km ou plus. À leurs yeux, cela signifie que la présence d’une grand-mère améliore le succès reproductif de ses enfants.
Toutefois, rien ne dit que l’entraide entre générations existait chez les hominiens où la
ménopause est apparue, avant qu’ils se dispersent à partir de l’Afrique. Par ail eurs, les conditions de vie de ces hommes n’ont sans doute pas grand-chose à voir avec cel es qui prévalaient naguère en Finlande et au Canada.
5.5. ORIGINE DE L'APOPTOSE
À première vue, l'existence de l'apoptose ne devrait pas poser trop de problèmes aux évolutionnistes, car el e favorise la reproduction des animaux en participant à la construction et à l'entretien du soma. Mais l'origine du phénomène reste mystérieuse. Certains chercheurs orientent leurs réflexions en se laissant guider par les rapports qui existent entre l’apoptose et la mitose. D’autres spéculent sur le rôle des mitochondries dans l’apparition du phénomène. 5.5.1. RELATION ENTRE L'APOPTOSE ET LA MITOSE
L'apoptose présente des similitudes avec la mitose, particulièrement en ce qui concerne les mécanismes de régulation. Par exemple, la protéine P53 des mammifères contrôle à la fois l'entrée en division et le déclenchement de l'apoptose. Ces ressemblances incitent à considérer l'apoptose comme une mitose dévoyée. Sur la voie qui la conduit à se diviser, toute cel ule serait donc forcée de choisir en permanence entre la vie et la mort.
Ce point de vue ne semble pas justifié, au moins pour deux raisons. En premier lieu, la
participation de la protéine P53 au contrôle de la mitose n'est pas un phénomène général, puisqu'el e ne s'observe ni chez C. elegans, ni chez la drosophile. C'est apparemment une innovation propre aux vertébrés. En second lieu, les cel ules en arrêt mitotique peuvent aussi se suicider, comme cela se produit notamment au cours de l'embryogenèse de C. elegans. Il n'y a donc pas de lien général entre la mitose et l'apoptose.
5.5.2. ROLE DES MITOCHONDRIES DANS L'APPARITION DE L'APOPTOSE
D'autres réflexions sont nées du fait que les mitochondries prennent souvent part à l'exécution de l'apoptose. Certains auteurs voient dans cette participation le vestige d'une guerre intestine qui aurait opposé les proto-mitochondries aux cel ules qu'el es essayaient d'envahir. À la longue, les partenaires ont fini par conclure la paix, à tel point qu'ils ne peuvent plus vivre les uns sans les autres, tout au moins dans les milieux riches en oxygène. Mais la paix est fragile et les hostilités peuvent reprendre lorsque les circonstances l'exigent et, en particulier, quand la cel ule a subi des lésions irréparables.
Cette proposition paraît judicieuse. Mais el e crée un problème de chronologie. Si l'apoptose est
réel ement la conséquence d'un conflit intérieur, el e a dû apparaître dès le début de l'évolution des eucaryotes et se maintenir pendant l'énorme interval e de temps (de l'ordre du mil iard d'années) qui a précédé l'apparition des animaux. On devrait donc retrouver chez les eucaryotes unicel ulaires des formes d'apoptose et peut-être des traces d’apoptosome.
Les ciliés* et les foraminifères* pratiquent une espèce d’apoptose. Au début du processus
sexuel, le ou les noyaux somatiques se détruisent, tandis que le ou les noyaux de nature germinale entrent en méiose (chapitre 1, section 1.2 et annexe II).
Les cel ules de levure possèdent des orthologues de certains facteurs pro-apoptotiques
identifiés chez les mammifères, tels que les protéines HTRA2 et AIF (annexe III.C.2). El es possèdent au moins une caspase, appelée YCA1 (Yeast Caspase 1). Quand el es meurent victimes de l’une et l’autre forme de sénescence (réplicative et chronologique), les cel ules de levure présentent certains signes caractéristiques de l’apoptose, et notamment une fragmentation rapide de la chromatine
Un phénomène semblable a été observé chez certains flagel és parasites appartenant au
groupe des trypanosomes quand ils subissent diverses agressions à un stade déterminé de leur cycle vital.
5.5.3. DIFFICULTÉS D'INTERPRÉTATION
Chez les ciliés, l’existence d’une forme d’apoptose nucléaire ne pose pas de problème évolutif sérieux. El e assure le renouvel ement du matériel génétique employé durant la phase de multiplication végétative qui précède le recours au processus sexuel (annexe II). Cette amputation a sans doute procuré un avantage sélectif aux cel ules qui l'ont adoptée, parce qu'el e leur a permis construire des noyaux et donc des cel ules de très grande tail e, mais incapables d'accomplir la méiose. Ces noyaux sont détruits lors de la conjugaison, puis reconstitués.
En revanche, on voit mal quel bénéfice peut procurer à la levure ainsi qu’aux trypanosomes la
mort qu'ils sont censés s'infliger, à moins de supposer que la disparition d'une partie des cel ules profite à cel es qui restent en vie. Mais cette interprétation recourt au concept d’altruisme* et de sélection de groupe*, dont il faut se méfier. Certains spécialistes de l’apoptose chez la levure s’efforcent de rester dans l’orthodoxie darwinienne. Ils font remarquer que dans les colonies vieil issantes poussant sur un milieu solide, ce sont les cel ules du centre qui meurent préférentiel ement. Ils assimilent ces communautés à des organismes multicel ulaires dont certains éléments entameraient une sorte de différenciation conduisant à la mort. Ce sacrifice bénéficierait à l’ensemble de la colonie, notamment parce qu’el e éviterait de consommer des substances nutritives indispensables à la survie des cel ules de la périphérie. Il s’agirait donc bien d’une sorte d’altruisme, difficile à admettre par les darwinistes. On voit que l’origine de l’apoptose pose des problèmes qui sont loin d’être résolus.
5.6. CONCLUSION La découverte des gérontogènes a déconcerté beaucoup de biologistes, qui ne concevaient pas que de tels gènes puissent exister, car leurs produits contribuent à écourter la vie des animaux, donc à compromettre leur succès reproductif. La découverte de l'apoptose en tant que mécanisme de mort « volontaire » a été moins dérangeante, parce qu'el e n'interfère pas directement avec la reproduction.
Mais les darwinistes peuvent être rassurés. En fait, la plupart des mutations qui accroissent de
manière notable la longévité des animaux ont pour eux des inconvénients qui se manifestent dès le début de leur vie. On peut donc penser que la sélection naturel e a pu consentir à ce que des gènes aient des effets néfastes, parce que leurs produits ont un effet bénéfique pour les individus avant qu'ils ne deviennent aptes à se reproduire.
BIBLIOGRAPHIE Systèmes anti-sénescence
Blackburn EH, Greider CW. Telomeres. Ouvrage col ectif. New York: Cold Spring Harbor Laboratory Press,
Chattopadhyay MK, Tabor CW, Tabor H. Polyamines protect Escherichia coli cel s from the toxic effect of
oxygen. Proc Natl Acad Sci USA 2003; 100: 2261-5
Guarente L. Sir2 and aging-the exception that proves the rule. Trends Genet 2001; 17: 391-2.
Lue NF, Jiang S. Reverse transcriptase at bacterial telomeres. Proc Natl Acad Sci USA 2004; 101: 14307-8 Altruisme
Aristote. Histoire des animaux. Traduction de Tricot J. Paris: Vrin, 1987.
Evans TA, Wal is EJ, Elgar MA. Making a meal of mother. Nature 1995; 376: 299.
Hamilton WD. Extraordinary sex ratios. Science 1967; 156: 477-88.
Mayr E. The objects of selection. Proc Natl Acad Sci USA 1997; 94: 2091-4.
Sigmund K, Hauert C. Altruism. Curr Biol 2002; 12: R270-2.
West SA, Gardner A, Griffin AS. Altruism. Curr Biol 2006; 16: R482-3. Théories du vieillissement
Flatt TF, Promislow DEL. Stil pondering an age-old question. Science 2007: 318: 1251256-7.
Haldane JBS. New paths in genetics. Londres: Allen et Unwin, 1941.
Kirkwood TBL.Understanding the old science of aging. Cell 2005; 120: 437-47.
Kirkwood TBL. Evolution of ageing. Nature 1977; 370: 301-4.
Kirkwood TBL, Austad SN. Why do we age? Nature 2000; 408; 233-8.
Medawar PB. An unsolved problem in biology. Londres: Lewis, 1952.
Reznick DN, Bryant MJ, Roff D, Ghalambor CK, Ghalambor DE. Effect of extrinsic mortality on the evolution of
senescence in guppies. Nature 2004; 431: 1095-9
Wal ace AR. Cité dans Weismann A. Essays upon heredity and kindred biological problems. Oxford: Clarendon
Wil iams GC. Pleiotropy, natural selection, and the evolution of senescence. Evolution 1957; 11: 398-411. Origine de la ménopause
Hawkes K. The grandmother effect. Nature 2004; 428: 128-9.
Lahdenperä M, Lummaa V, Hel e S, Tremblay M, Russel AF. Fitness benefits of prolonged post-reproductive
lifespain in women. Nature 2004; 428: 178-81.
Packer C, Tatar M, Col ins A; Reproduction cessation in female mammals. Nature 1998; 392: 807-11.
Shanley DP, Kirkwood TBL. Evolution of the human menopause. BioEssays 2001; 23: 282-7.
Sherman PW. The evolution of menopause. Nature 1998; 392; 759-61.
Thomson ME, Jones JH, Pusey AE, Brewer-Marsden S et al. Aging and fertility patterns in wild chimpanzees
provide insights into the evolution of menopause. Curr Biol 2007; 17: 2150-6. Origine de l’apoptose
Ameisen JC. La sculpture du vivant. Paris: Seuil, 1999.
Ameisen JC, Idziorek T, Bil aut-Mulot O, Loyens M, Tissier J-P, Potentier A, Ouaissi A. Apoptosis in a unicel ular
eukaryote (Trypanosoma cruzi): implications for the evolutionary origin and role of programmed cel death in the control of cel proliferation, differentiation and survival. Death Differ 1995; 2: 285-300.
Blackstone NW, Green DR. The evolution of a mechanism of cel suicide. BioEssays 1999; 21: 84-8.
Büttner S, Eisenberg T, Herker E, Carmona-Gutierrez D et al. Why yeasts cel s can undergo apoptosis: death
times of peace, love, and war. J Cell Biol 2006; 175: 521-5.
Lawen A. Apoptosis-an introduction. BioEssays 2003; 25: 888-96.
Melino G. The sirens' song. Nature 2001; 412: 23.
Welburn SC, Dale C, Ellis D, Beecroft R, Pearson TW. Apoptosis in procyclic Trypanosoma brucei rhodesiense in vitro. Death Differ 1996; 3: 229-36.
Wysocki R, Kron SJ. Yeast cel death during DNA damage arrest is independent of caspase or reactive oxygen
species. J Cell Biol 2004; 166: 311-6.
that one treatment per week will be appropriate for most patients, however prescribers may exercise their clinical judgement should a patient request more than this. Drugs for ED may have a street value and the issue of diversion should be borne in mind when considering prescription quantities. February 2009 OSTEOPOROSIS GUIDELINES: Alendronate is the drug of first choice on DRUG
Curriculum Rationale The ECLC curriculum will meet the needs of the children in all aspects of their development. The project approach for curriculum planning will ensure the program is child initiated and inquiry based. The ECLC program will be appropriate to the age of the children and encourage their intellectual, emotional, social, creative and physical growth. The ECLC curriculum is a
 5.3.3. LES THÉORIES MODERNES DU VIEILLISSEMENT
5.3.3. LES THÉORIES MODERNES DU VIEILLISSEMENT 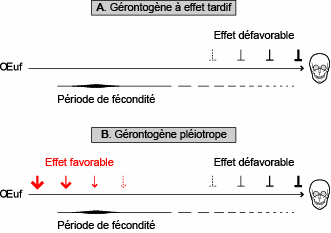 diminuant la vitalité les individus devenus vieux, pourvu qu'el es augmentent les chances de survie des jeunes. Suivant ce point de vue, la longévité des animaux serait fixée par un subtil équilibre entre les effets contradictoires d'un certain nombre de gérontogènes.
Fig. 5.1. Modes d'action des gérontogènes.
diminuant la vitalité les individus devenus vieux, pourvu qu'el es augmentent les chances de survie des jeunes. Suivant ce point de vue, la longévité des animaux serait fixée par un subtil équilibre entre les effets contradictoires d'un certain nombre de gérontogènes.
Fig. 5.1. Modes d'action des gérontogènes. 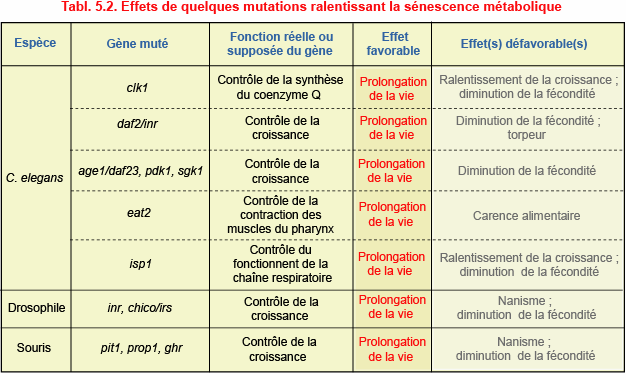 5.3.9. DOUBLE EFFET DES FACTEURS APTES À PROLONGER LA VIE
5.3.9. DOUBLE EFFET DES FACTEURS APTES À PROLONGER LA VIE