Kidney International, Vol. 65 (2004), pp. 531–539
ION CHANNELS – MEMBRANE TRANSPORT – INTEGRATIVE PHYSIOLOGY
Regulation of gene expression by dietary Ca2+ in kidneysof 25-hydroxyvitamin D3-1a-hydroxylase knockout mice
JOOST G.J. HOENDEROP, HELENA CHON, DIMITRA GKIKA, HANS A.R. BLUYSSEN, FRANK C.P. HOLSTEGE, RENE ST-ARNAUD, BRANKO BRAAM, and RENE J.M. BINDELS Department of Physiology, University Medical Centre Nijmegen, Nijmegen, The Netherlands; Department of Nephrology andHypertension and Genomics Laboratory, Department of Physiological Chemistry, UMC Utrecht, The Netherlands; and GeneticsUnit, Shriners Hospital for Children, Montreal, CanadaRegulation of gene expression by dietary Ca2+ in kidneys of
Vitamin D is a major regulator of Ca2+ and phos-
25-hydroxyvitamin d3-1a-hydroxylase knockout mice.
phate homeostasis and is essential for proper devel-
Background. Pseudovitamin D deficiency rickets (PDDR)
opment and maintenance of bone. The active form of
is an autosomal disease, characterized by undetectable levelsof 1,25-dihydroxyvitamin D
vitamin D, 1a,25-dihydroxyvitamin D3 (1,25(OH)2D3) is
ondary hyperparathyroidism. Mice in which the 25-hydroxy-
synthesized from its precursor 25-hydroxyvitamin D3 by
vitamin D3-1a-hydroxylase (1a-OHase) gene was inactivated,
the renal cytochrome P450 enzyme 25-hydroxyvitamin
presented the same clinical phenotype as patients with PDDR.
D3-1a-hydroxylase (1a-OHase). The importance of this
Methods. cDNA Microarray technology was used on kidneys
enzyme is reflected by severe disorders resulting from
of 1a-OHase knockout mice to study the expression profile ofrenal genes in this Ca2+-related disorder. Genome wide molec-
mutations identified in the gene encoding 1a-Ohase, in-
ular events that occur during the rescue of these mice by high
cluding pseudovitamin D-deficiency rickets (PDDR) also
dietary Ca2+ intake were studied by the use of 15K cDNA mi-
known as vitamin D-dependent rickets type I (VDDR-
I). Recently, the genetic association of 1a-OHase with
Results. 1a-OHase knockout mice fed a normal Ca2+ diet
PDDR has been confirmed by inactivation of the 1a-
developed severe hypocalcemia, rickets and died with an aver-age life span of 12 ± 2 weeks. Intriguingly, 1a-OHase−/− mice
OHase gene in mice [1, 2]. Two laboratories indepen-
supplemented with an enriched Ca2+ diet were normocalcemic
dently generated 1a-OHase knockout mice strains that
and not significantly different from wild-type mice. Inactivation
represent valuable animal models for PDDR since they
of the 1a-OHase gene resulted in a significant regulation of ±
display undetectable 1,25(OH)2D3 levels, hypocalcemia,
1000 genes, whereas dietary Ca2+ supplementation of the 1a-
secondary hyperparathyroidism, and failure to thrive
OHase−/− mice revealed ± 2000 controlled genes. Interestingly,557 transcripts were regulated in both situations implicating the
[1, 2]. The 1a-OHase knockout mice (1a-OHase−/−)
involvement in the dietary Ca2+-mediated rescue mechanism of
developed distinct histologic evidence of rickets and
the 1a-OHase−/− mice. Conspicuous regulated genes encoded
for signaling molecules like the PDZ-domain containing pro-
The absence of vitamin D is expected to cause
tein channel interacting protein, FK binding protein type 4,
widespread changes in gene expression in the kidney. On
kinases, and importantly Ca2+ transporting proteins includingthe Na+-Ca2+ exchanger, calbindin-D
one hand, vitamin D receptor (VDR) activation will be
decreased resulting in altered gene expression. On the
Conclusion. Dietary Ca2+ intake normalized disturbances in
other hand, Ca2+ sensing by the Ca2+ sensing receptor
the Ca2+ homeostasis due to vitamin D deficiency that were
(CaSR) will be changed due to the hypocalcemia in 1a-
accompanied by the regulation of a subset of renal genes, in-
OHase mice. The importance of VDR activation versus
cluding well-known renal Ca2+ transport protein genes, but alsogenes not previously identified as playing a role in renal Ca2+
CaSR activation has recently been challenged by the ob-
servation that many of the phenotypic effects of VDRknockout animals can be prevented by a “rescue diet”with lactose, Ca2+, and phosphate [3–5]. This observationis supportive for the view that vitamin D becomes rele-
Key words: TRPV5, ECaC1, vitamin D, calcium reabsorption, PDDR.
vant at the point where an organism is denied an abundantCa2+ supply.
In a previous study, we have found indications for these
and in revised form August 27, 2003Accepted for publication September 10, 2003
Ca2+-related actions in the 1a-OHase−/− knockout mice[6]. This study showed that high dietary Ca2+ intake nor-
C 2004 by the International Society of Nephrology
malizes the expression levels in 1a-OHase−/− of the Ca2+
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
transport proteins, including the epithelial Ca2+ channel
Reverse transcription/labeling
(TRPV5), calbindin-D28K, Na+-Ca2+ exchanger (NCX1),
Samples were pooled per group in equal amounts
and plasma membrane adenosine triphosphatase (AT-
of total RNA per subject. For each reaction, 5 lg
Pase) (PMCA1b), and contributes in this way to the nor-
of total RNA was annealed to 1 lg of Oligo-dT12−18
malization of blood Ca2+ levels [6].
primer (Invitrogen Life Sciences) at 70◦C for 15 min-
The aim of the present study was to identify new
utes. Then, reverse transcription (RT) was performed in
genes important for the maintenance of the Ca2+ bal-
a 30 lL reaction containing 200 U SuperScript II re-
ance. The hypothesis is that vitamin D deficiency results
verse transcriptase (Invitrogen Life Sciences), 2 nmol
in widespread gene expression alterations in the kidney,
deoxyguanosine triphosphate (dGTP), deoxyadenosine
which are in part due to diminished CaSR activation by
triphosphate (dATP), and deoxycytidimine triphosphate
decreased plasma Ca2+ concentrations. Furthermore, we
(dCTP), 0.5 nmol deoxydithiothreitol (dTTP) (Amer-
hypothesized that genes, differentially expressed in 1a-
sham Biosciences Corp., Biscataway, NJ, USA), 1.5 nmol
OHase−/− mice, could be rescued by dietary Ca2+ via
of aminoallyl-deoxyuridine triphosphate (dUTP) (Amer-
a vitamin D–independent mechanism. To address these
sham Biosciences Corp.) at 42◦C for 2 hours. After
questions, DNA microarray analysis of gene expression
2 minutes at 95◦C, a 20 lL mixture of 1 mol/L NaOH
was compared between 1a-OHase−/− and 1a-OHase+/−
(Riedel-de Ha ¨en, Seelze, Germany) and 0.5 mol/L of
mice supplemented with dietary Ca2+.
ethylenediaminetetraacetic acid (EDTA), pH 8.0 (SigmaChemical Co., St. Louis, MO, USA) were added and incu-bated at 65◦C for 30 minutes to hydrolyze RNA. The reac-
tion was neutralized by adding 25 lL of 1 mol/L of Hepes,
Animal protocol
pH 7.5 (Sigma Chemical Co.). Samples were purified onMicrocon-30 columns (Millipore, Billerica, MA, USA)
25-hydroxyvitamin D3-1a-hydroxylase knockout mice
and reduced to a volume of 8 lL. Cy3 and Cy5 mono-
were recently generated by targeted ablation of exon
functional reactive dyes (Amersham Biosciences UK
8 encoding the HEME binding domain of the en-
Unlimited, Buckinghamshire, UK) dissolved in dimethyl
zyme [1]. 1a-OHase knockout mice were genotyped
sulfoxide (DMSO) (Riedel-de Ha ¨en) and 1 lL of
by Southern blot analysis at an age of 3 weeks di-
0.5 mol/L sodium bicarbonate (pH 9.0; Sigma, St Louis,
rectly after the weaning period as described previously
MO) were added to the cDNA samples and incubated at
[1]. Initial characterization of the 1a-OHase knockout
room temperature for 1 hour. Dyes were quenched by in-
mice demonstrated that there are no significant dif-
cubation with 4.5 lL of 4 mol/L Hydroxylamine (Aldrich
ferences between wild-type (1a-OHase+/+) and het-
Chemicals, Co., Milwaukee, MI, USA) at room temper-
erozygous 1a-OHase knockout mice (1a-OHase+/−) [1].
ature for 15 minutes. Labeled samples were purified on
The heterozygous were, therefore, used as control an-
Chromaspin-30 columns (BD Biosciences Clontech, Palo
imals. 1a-OHase+/− and homozygous 1a-OHase (1a-
Alto, CA, USA). Samples from 1a-OHase−/− mice and
OHase−/−) knockout mice were fed from weeks 3 to 8
1a-OHase+/− mice with Ca2+ supplementation were la-
either a normal diet [1.1% (wt/wt) Ca2+, 0.8% (wt/wt)
beled with Cy3 and samples from 1a-OHase+/− mice and
phosphorus, and 0% (wt/wt) lactose], a Ca2+-enriched
1a-OHase−/− mice with Ca2+ supplementation were la-
diet [2% (wt/wt) Ca2+, 1.25% (wt/wt) phosphorus, and
20% (wt/wt) lactose] (Harlan Tekled, Wisconsin, MA,USA). Animals (N = 6 in each group) were sacrificedat an age of 8 weeks and blood and kidney samples
Hybridization of cDNA chips
were taken. The Animal Ethics Board of the Univer-
Mouse NIH 15K cDNA chips (Ontario Microarray
sity of Nijmegen (Nijmegen) and Shriners Hospital for
Centre, Toronto University, Canada) were prehybridized
Children (Montreal) approved all animal experimental
for at least 45 minutes in a filtered (0.22 micron Minisart-
plus filter, Sartorius AG, G ¨ottingen, Germany) andpreheated buffer (37◦C) containing 1 g bovine serum al-bumin (BSA) (Sigma Chemical Co.), 25 mL 20× standard
RNA isolation
sodium citrate (SSC), 25 mL pure formamide (Merck,
RNA was extracted from whole kidney from four sub-
Darmstadt, Germany), 1 mL 10% (wt/vol) sodium dode-
jects per group using the TRIzol reagent using the pro-
cyl sulfate (SDS) (Bio-Rad Laboratories, Hercules, CA,
cedure recommended by the supplier (Invitrogen Life
USA), in a total volume of 100 mL. After washing the
Sciences, Carlsbad, CA, USA). The quality and quantity
slides with distilled water and isopropanol (Mallinkrodt
of the RNA samples was determined with the Bioana-
Baker BV, Deventer, The Netherlands), slides were spun
lyzer (Agilent, Palo Alto, CA, USA) using the Eukaryote
dry. Slides were then loaded with 100 to 150 ng labeled
cDNA in a filtered (0.22 micron) and preheated buffer
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
containing 25% (wt/vol) formamide (Bio-Rad Labora-tories), 5× SSC [sodium chloride) (Riedel-de Ha ¨en)];
sodium citrate dihydrate (Merck, Dramstadt, Germany);
0.1% (wt/vol) SDS; 200 lg/mL of Herring Sperm DNA(Invitrogen Life Sciences); and tRNA (Roche Molecular
Diagnostics GmbH, Mannheim, Germany) that had beendenatured at 95◦C for 5 minutes and incubated at 42◦C
overnight in acclimatized hybridization chambers (Corn-
ing Incorporated Life Sciences, Acton, MA, USA). Slides
were then washed first, in 100 mL 1× SSC with 0.2%(wt/vol) SDS; second, in 0.1× SSC with 0.2% (wt/vol)
Scanning/quantification of cDNA chips
Slides were scanned on the ScanArray 4000XL
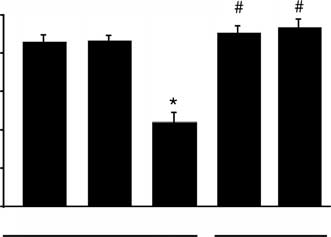
Fig. 1. The effect high dietary Ca2+ intake on the serum Ca2+ concen-
(BioDiscovery, Marina del Rey, CA, USA) at laser in-
tration in 1a-hydroxylase (1a-OHase−/−) mice. Values are presented
tensities of 85 to 90 and photomultiplier tube sensitivity
as means ± SEM (N = 6). ∗P < 0.05, significant different from 1a-OHase+/+ and 1a-OHase+/− mice fed a normal Ca2+ diet containing
of 60 to 65 at a resolution of 10 lm. Images were quanti-
1.1% (wt/vol) Ca2+; #P < 0.05, significant different from 1a-OHase−/−
fied using Imagine software (BioDiscovery). Raw mean
mice on normal Ca2+ diet containing 1.1% (wt/wt) Ca2+.
intensity data were averaged per duplicate spot and nor-malized on median ratios on the chip by correcting the
analysis of variance (ANOVA). In the case of significance
Cy5 values. Spots with both Cy3 and Cy5 intensities below
(P < 0.05), individual groups were compared by contrast
background plus two times the standard deviation of the
background signal and spots with intensities exceeding45000 were excluded from further analysis. Genes were
considered significantly regulated when log2 transformed
Phenotypical characterization of the 1a-OHase−/− mice
Inactivation of the 1a-OHase gene in mice resulted
Clustering analysis and gene ontology annotations
in severe hypocalcemia with serum Ca2+ concentrations
Hierarchical clustering of microarray data was per-
around 1.2 mmol/L, whereas 1a-OHase+/− mice, which
formed using the expression profiler tool EPCLUST (Eu-
are not distinguishable from 1a-OHase+/+ mice, dis-
ropean Bioinformatics Institute; Jaak Vilo, Cambridge,
played normal plasma Ca2+ concentrations (Fig. 1). 1a
UK). Average linkage (average distance) clustering
-OHase−/− mice fed a regular Ca2+ diet containing 1.1%
based on correlation measure-based distance was
(wt/wt) Ca2+ developed rickets and died with an aver-
performed on data for which ratios were >0.7 or <−0.7 in
age life span of 12 ± 2 weeks (data not shown). Inter-
at least one comparison. Ratios of the genes were coupled
estingly, supplementation of these mice with an enriched
to the biologic processes classification of the Gene Ontol-
Ca2+ diet containing 2% (wt/wt) Ca2+ normalized plasma
ogy Consortium (GO Consortium) [7] using proprietary
Ca2+ concentrations that were not significantly different
software and Access 2000 (Microsoft Corporation, Red-
from 1a-OHase control mice (Fig. 1). This latter diet re-
mond, WA, USA). For each biologic process category,
sulted in a normal development and life span of these
the total number of annotated genes and the percentage
of significantly changed genes were counted, and the av-
Gene expression profiles of 1a-OHase−/− mice
erage ratio was calculated. Interesting categories were
compared with 1a-OHase+/− mice
defined as groups containing at least five genes and thathad either a mean overall ratio of >0.3 or <0.3 or had at
Around 1000 genes were differently regulated in 1a
least 20% of the genes of that group changed significantly.
-OHase−/− mice versus the 1a-OHase+/− mice, whereasroughly 2000 genes were significantly regulated by di-
Serum Ca2+ measurements
etary Ca2+ in the vitamin D–deficient mice. Interestingly,557 transcripts, that account for 4% of the total num-
Serum Ca2+ concentrations were measured using a col-
ber of 15,000 scanned genes, were regulated under both
orimetric assay kit as described previously [8].
conditions. This is demonstrated in the Venn diagramsin Figure 2A. Moreover, only minor changes in ex-
Statistical analysis
pression were induced by dietary Ca2+ in 1a-OHase+/−
The serum Ca2+ data were expressed as the mean ±
mice. Figure 2B and C shows the numbers of up- and
SEM. Overall statistical significance was determined by
down-regulated genes in all the experimental conditions
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Shared −/− vs. −/− Shared +/− vs. +/−
Fig. 2. Number of genes regulated when comparing the three condi- tions 1a-hydroxylase (1a-OHase−/−) mice versus 1a-OHase+/− mice (left), 1a-OHase−/− mice versus 1a-OHase−/− mice supplemented Fig. 3. Hierarchical clustering comparisons with dietary Ca2+ (middle) and 1a-OHase+/− mice versus Ca2+ sup- hydroxylase (1a-OHase−/−) mice versus 1a-OHase+/− mice, 1a plemented 1a-OHase+/− mice (right). The number of total-regulated -OHase−/− mice versus 1a-OHase−/− mice treated with Ca2+ and
(A), up-regulated (B), or down-regulated (C) transcripts on the DNA
1a-OHase+/− mice compared to 1a-OHase+/− mice fed the enriched
chip are depicted. Genes are shown in the overlapping regions that areregulated in two conditions. Ca2+ diet]. Genes that are significantly up- (dark) or down-regulated (light) are shown.
and includes an overview of the gene expression profileof each condition separately and in combination.
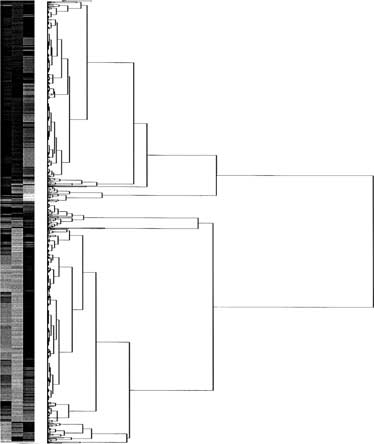
Next, we performed a hierarchical clustering of the
2 above 0.7 and below –0.7) that were inversely
regulated as a consequence of 1a-OHase gene inactiva-
three comparisons in the present study: 1a-OHase−/− ver-
tion (Fig. 4A) and dietary Ca2+ supplementation in the
sus 1a-OHase+/−, 1a-OHase−/− versus 1a-OHase−/− sup-
vitamin D–deficient state (Fig. 4B). Genes depicted in yel-
plemented with Ca2+ and 1a-OHase+/− compared to 1a
low were regulated in a similar direction in the described
-OHase+/− treated with Ca2+ (Fig. 3). A large portion of
the expression profiles of knockout versus 1a-OHase+/−and 1a-OHase−/− versus 1a-OHase−/− treated with Ca2+were similarly regulated, but there were also some genes
Gene expression changes specifically related
differentially regulated. The comparison of 1a-OHase−/−
to Ca2+ supplementation
mice versus 1a-OHase−/− mice treated with Ca2+ and 1a
Further analysis of the genes that were inversely regu-
-OHase+/− mice compared to 1a-OHase+/− mice supple-
lated by dietary Ca2+ supplementation and 1,25(OH)2D3
mented with Ca2+ yielded far less similarity, but a distinct
deficiency provides information about interesting can-
didates involved in the 1,25(OH)2D3-independent res-
Figure 4 shows representative differential expres-
cue mechanism by dietary Ca2+ (Table 1) (the com-
sion of 1a-OHase−/− versus 1a-OHase+/− mice in an
plete data set of the regulated genes is accessible
intensity-dependent fashion with Cy3/Cy5 scatter plots.
via http://www.genomics.med.uu.nl/pub/bb/kidney/). Two
Up-regulated and down-regulated genes formed two dis-
important cDNA transcripts, known to encode for Ca2+
tinct populations that clearly differentiate from the non-
transporting proteins in the distal part of the nephron,
regulated genes. Of interest is the group of genes (de-
calbindin-D28K and the Na+-Ca2+ exchanger (NCX1),
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout miceAnalysis using GO Consortium functional classification of genes
Differentially regulated genes were grouped into func-
tional profiles using GO Consortium annotation. In
both the comparison of the 1a-OHase−/− with the 1a-
OHase+/− mice and the comparison of the 1a-OHase−/−
mice with the 1a-OHase−/− mice receiving the high Ca2+
diet, alterations in some functional categories could be
identified. Of all 307 categories of biologic processes, 109
had five or more genes that were represented on the
Fig. 4. Differential expression patterns depicted in an intensity-
cDNA chip. Of these, 14 processes were considered to be
dependent fashion using Cy3/Cy5 scatter plots. Regulated genes by
changed in the 1a-OHase−/− mice versus 1a-OHase+/−
inactivation of the 1a-hydroxylase (1a-Ohase) gene (A) and supplemen-
mice and 32 processes in 1a-OHase−/− mice versus 1a-
tation of dietary Ca2+ in the vitamin D–deficient state (B) form twodistinct populations of genes. Genes that are regulated in two condi-
OHase−/− mice supplemented with Ca2+ comparison.
tions (when 1a-OHase+/− mice are compared with 1a-OHase−/− mice
Eleven categories were overlapping in both comparisons
and when 1a-OHase−/− supplemented with dietary Ca2+) are shown
(Fig. 5). Inactivation of the 1a-OHase gene and sup-
above the 0.7 log2 line (up-regulated in both comparisons) and belowthe −0.7 log
plementation of Ca2+ in 1a-OHase−/− mice resulted in
2 line (down-regulated in both comparisons). Genes that
are regulated in opposite directions in both comparisons are shown as
a regulation of genes involved in protein biosynthesis.
Remarkably, a subset of the biologic processes were reg-ulated in a similar fashion in both comparisons. Biologicprocesses such as glycolysis, peroxidase reaction, hydro-
were down-regulated in the 1a-OHase−/− mice and up-
gen transport, tricarbolic acid cycle, iron homeostasis,
regulated by dietary Ca2+ in the 1,25(OH)2D3-deficient
and lymph gland development were decreased in 1a-
animal. Previously, these genes were analyzed by quanti-
OHase−/− mice and reactivated through dietary Ca2+
tative RT-polymerase chain reaction (PCR) in the same
supplementation. Genes involved in muscle development
animal model and were found to be regulated in a similar
and induction of apoptosis were slightly up-regulated due
manner [6]. Analysis of other genes that were significantly
to the absence of 1,25(OH)2D3, which were not affected
regulated revealed several genes previously unknown to
by Ca2+ supplementation of the 1a-OHase−/− mice.
be involved in the Ca2+ homeostasis. New candidates in-clude the Ca2+-related genes like FK506 binding protein4, calmodulin, and several ion transporters (i.e., the K+
DISCUSSION
channel protein Kcnq1, the Na+-H+ exchanger, and the
In the present study, we demonstrated that dietary
voltage-dependent Ca2+ channel a1 subunit). Several ki-
Ca2+ intake is an important regulator of the Ca2+ home-
nases and kinase substrates controlling distinct signalling
ostasis in vitamin D–deficient 1a-OHase−/− mice, ulti-
pathways were regulated by dietary Ca2+ as well as a
mately normalizing blood Ca2+ levels. Microarray anal-
channel-interacting PDZ domain protein (Table 1).
ysis revealed that high dietary Ca2+ intake restores the
Additional analysis was performed to study genes that
disturbed genetic profile observed in 1a-OHase−/− mice,
followed the Ca2+ availability in 1a-OHase−/− and 1a
which is accompanied by a functional rescue of these
-OHase+/− mice. Although 1a-OHase+/− mice did not
1,25(OH)2D3-deficient animals. Gene expression analy-
show elevated levels of serum Ca2+ (Fig. 1), several
sis showed simultaneous regulation of genes with poten-
genes were found to be down-regulated in 1a-OHase−/−
tial importance for Ca2+ handling that were not previ-
mice without dietary supplementation and up-regulated
ously identified as such and a subset of genes that were
when 1a-OHase+/− mice were supplemented with Ca2+
regulated by dietary Ca2+ independently of vitamin D3.
(Table 2). These genes are thought to be interest-
Recently, Dardenne et al [1] generated an 1a-OHase
ing candidates that are controlled by Ca2+ irrespective
knockout strain that represents a valuable animal model
of the vitamin D state of the animal. Among them
for PDDR since these mice display all the clinical symp-
were genes involved in mitogen-activated protein kinase
toms observed in patients, including hypocalcemia and
(MAPK)/tyrosine kinase signaling and genes that are re-
rickets. These pathologic symptoms in the 1a-OHase−/−
sponsive to intracellular Ca2+ concentrations (Table 2).
mice resulted in an early death with an average life span
Subsequently, genes regulated by the 1a-OHase gene in-
of 12 weeks. Normalization of the plasma Ca2+ concen-
activation, but not sensitive to dietary Ca2+ supplemen-
trations by dietary Ca2+ supplementation was associ-
tation, were identified and depicted in Table 3. Typical
ated with a functional rescue of the 1a-OHase−/− mice
genes regulated by vitamin D only were identified in sev-
that made the recovered mice phenotypically undistin-
eral function groups, including Ca2+ transport proteins
guishable from wild-type mice. A previous study indi-
cated that the Ca2+ transport proteins, including TRPV5
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout miceTable 1. Summary of candidate genes identified using cDNA microarrays
v-crk-associated tyrosine kinase substrate
Depicted genes are reversely regulated by inactivation of the 1a-hydroxylase (1a-OHase) gene and supplementation by dietary Ca2+ in the vitamin D–deficient state.
The log2 ratios represent the change in expression when 1a-OHase−/− mice are compared to 1a-OHase+/− mice and when 1a-OHase−/− mice are supplemented withCa2+. The complete data set of the regulated genes is accessible via http://www.genomics.med.uu.nl/pub/bb/kidney/.
(previously named ECaC1), calbindin-D28K, and NCX1,
pendent of parathyroid hormone (PTH) and vitamin D.
were down-regulated in the 1a-OHase mice [6]. Interest-
This significant effect of hypocalcemia could indicate a
ingly, the expression of these proteins was normalized by
decrease in CaSR activation. The CaSR is an important
gateway by which extracellular Ca2+ can affect a variety
The present analysis demonstrated that known Ca2+
of second messenger systems, such as the phopsholipase
transport proteins represented on the cDNA microarray,
A2 (PLA2) and phopsholipase C (PLC), adenylate cy-
including calbindin-D28K and NCX1, were regulated by
clase, and MAPKs. The downstream actions of CaSR on
dietary Ca2+. Dietary Ca2+ supplementation in the 1a-
gene expression involve a large number of cellular pro-
OHase−/− mice had a maximum effect on NCX1 expres-
cesses, most notably secretory pathways of hormones,
sion suggesting that this basolateral extrusion protein
channels, and transporters (e.g., the thiazide-sensitive
is an important mechanism in the process of transcel-
NaCl cotransporter, K+ channels and aquaporin-2) and
lular Ca2+ reabsorption. In line with these findings are
apoptosis-related genes. In various clinical conditions as-
functional studies in cell model systems demonstrating
sociated with a disturbed Ca2+ homeostasis, vitamin D
that basolateral extrusion of Ca2+ is mainly achieved by
analogues are administered. The treatment of choice for
NCX1 [9, 10]. Interestingly, calbindin-D9K was not reg-
PDDR and for patients with chronic renal failure is long-
ulated by dietary Ca2+ supplementation. The expression
term replacement therapy with 1,25(OH)2D3. Notably,
of this Ca2+ binding protein was solely controlled by vi-
the currently applied strategy of vitamin D and Ca2+ sup-
tamin D in line with previous studies [6].
plementation to patients with chronic renal failure has
Dardenne et al [11] investigated recently bone histol-
been associated with undesirable effects, such as vascu-
ogy and histomorphometry in 1a-OHase−/− mice sup-
lar calcification and calciphylaxis. It would be interesting
plemented with Ca2+ confirming that the rickets and
to compare the normalization of plasma Ca2+ levels by
osteomalacia were prevented. The Ca2+ rescue diet also
Ca2+ supplementation with the treatment with vitamin D
restored the biomechanical properties of the bone tis-
sue within normal parameters. Blood biochemical anal-
Hierarchical cluster analysis of the 15K cDNAs demon-
ysis revealed that the Ca2+ diet corrected the secondary
strated a remarkable overlap in the expression profiles
hyperparathyroidism. This points at an important role
between inactivation of the 1a-OHase gene compared to
for the decreased prevailing calcium levels per se, inde-
1a-OHase−/− mice and supplementation of dietary Ca2+
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout miceTable 2. List of genes that are regulated by the Ca2+ availability in 1a-hydroxylase (1a-OHase−/−) and 1a-OHase+/− mice with their
The log2 ratios represent the change in expression when supplemented with Ca2+ in 1a-OHase−/− mice and in 1a-OHase+/− mice. Table 3. List of genes that are significantly regulated by inactivation of the 1a-hydroxylase (1a-Ohase) gene and not by dietary Ca2+
supplementation with their corresponding ratios
ATPase H+-transporting lysosomal proton pump
ATP binding cassette sub-family C (CFTR/MRP)
K+ small conductance Ca2+ activated channel
The log2 ratios represent the change in expression when 1a-OHase−/− mice are compared with 1a-OHase+/− and 1a-OHase−/− mice versus 1a-OHase−/− mice
in 1a-OHase−/−, whereas only a minor overlap was ob-
PDZ domain protein (CIPP), FK506 binding protein 4,
served between 1a-OHase−/− with 1a-OHase+/− mice.
Interestingly, several transcripts, unknown to be involved
CIPP was found to interact with several ion chan-
in Ca2+ homeostasis, were significantly regulated. Pre-
nels including the acidic sensing ion channel ASIC3 and
vious studies from our group have shown that TRPV5,
the inward rectifying potassium channel Kir [13, 14].
not present on the microarray chip, facilitates the rate-
CIPP is a PDZ domain-containing protein that con-
limiting step in the process of transcellular Ca2+ trans-
tains four PDZ domains. Interestingly, TRPV5 contains
port [12]. With respect to the gatekeeper function of this
both in the amino and carboxyl-termini type I (S/T-X-
apical Ca2+ influx channel, it will be interesting to discuss
V/L) PDZ-interacting domains, that are similar to the
three regulated cDNAs encoding a channel-interacting
carboxyl-terminal sequence of the ASIC3 protein [15]. Hoenderop et al: Gene profiling of 1a-hydroxylase knockout miceFig. 5. A subset of regulated functional gene groups based on Gene Ontology (GO) with differential regulation due to 1a-hydroxylase (1a-OHase) inactivation or Ca2+ supplemen- tation. Displayed GO groups contained at
least five genes, had either a mean overall ra-tio of at least 0.3 or had at least 20% of the
−1.5 −1.0 −0.5 0.0 0.5 1.0 1.5−1.5 −1.0 −0.5 0.0 0.5 1.0 1.5
genes changed significantly in both compar-isons. Box plots indicate the quartiles of the
PDZ-containing proteins can indeed influence the cell
of the TRP family [25, 26]. Niemeyer et al [26] identified
surface expression of ion channel proteins by, for in-
a new calmodulin binding site in the carboxyl-terminus
stance, affecting their insertion, endocytosis, and recy-
of the TRPV5 homolog TRPV6. These investigators
showed that Ca2+-dependent calmodulin-binding to
The second transcript, FK506 binding protein 4
TRPV6 facilitates channel inactivation, which was coun-
(FKBP4), has been implicated as an important reg-
teracted by protein kinase C (PKC)-mediated phospho-
ulation of ion channel activity. Of interest, the
rylation of this putative calmodulin binding site. Based
transient receptor potential-like channel (TRPL), a
on the observed genomic regulation of calmodulin it will
Ca2+-permeable cation channel found in Drosophila pho-
be of interest to investigate whether this Ca2+ sensor
toreceptor cells that shares 25% homology and an iden-
interacts with TRPV5. However, functional studies are
tical structural topology with TRPV5, is modulated by
required to study the role of calmodulin expression in reg-
FKBP4 [18]. Intriguingly, renal transplant recipients re-
ulating the Ca2+ balance and in particular the activity of
ceiving the immunosuppressive drug FK506 (also known
TRPV5. The afore-mentioned genes were reversely reg-
as tacrolimus), which is the substrate for FKBP4, had
ulated by inactivation of the 1a-OHase gene and normal-
strongly reduced renal calbindin-D28K protein levels and
ized in expression by dietary Ca2+, which raises the attrac-
increased urine Ca2+ excretion [19, 20]. Furthermore, the
tive hypothesis that these putative candidates regulate
treatment caused intratubular calcification [21]. The data
Ca2+ reabsorption in general and in particular TRPV5
suggest a link between the observed regulation of FKBP4
activity, and thereby, restoring the hypocalcemia and
in the present study and the molecular regulation of Ca2+
subsequently other pathologic symptoms in the vitamin
reabsorption in the distal part of the nephron.
The third interesting candidate regulated on the chip
In addition to the previously discussed reversely regu-
was calmodulin. Calmodulin is a major cellular sensor
lated genes, a subgroup of genes was regulated by dietary
of Ca2+ signalling and interacts with numerous proteins
Ca2+ independent of the vitamin D status. These genes
associated with cellular second messenger systems [22,
were found to be up- or down-regulated by Ca2+ in 1a
23]. Changes in intracellular Ca2+ concentration regulate
-OHase+/− and 1a-OHase−/−. An interesting candidate
calmodulin in various distinct ways. First, at the cellular
is MAPK (p70/p85 s6 kinase), an enzyme that has been
level, by directing its subcellular distribution. Second, at
implicated in many cellular signaling processes, including
the molecular level, by promoting different modes of as-
voltage-operated Ca2+ channels [27]. Furthermore, cD-
sociation with many target proteins. Third, by directing
NAs encoding proteins, which are involved in activation
a variety of conformational states in calmodulin that re-
of MAPK, are coregulated like syntrophin. Anchoring of
sult in target-specific activation. Fourth, the present study
ion channels at specific subcellular sites is critical for sig-
suggests an association between serum Ca2+ levels and
naling, but the mechanisms underlying channel localiza-
the expression of calmodulin. Calmodulin -dependent
tion and clustering are largely unknown. Molecules such
regulation of protein kinases illustrates a potential mech-
as ankyrin and syntrophin, that bind ion channels, may
anism by which Ca2+-sensing proteins can recognize and
be important to maintain a high channel density at the
generate affinity as well as specificity for effectors in a
plasma membrane [28]. Gene regulation was studied in
Ca2+-dependent manner. It has been demonstrated that
total kidney RNA, although active Ca2+ reabsorption is
calmodulin interacts with several Ca2+ channels, includ-
restricted to the distal part of the nephron. The observed
ing voltage-gated Ca2+ channels [24], but also members
genomic regulation is, therefore, an averaged mRNA
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
difference measured in the whole kidney and studies with
dihydroxyvitamin D3 in 25-hydroxyvitamin D3–1a-hydroxylase
isolated tubules or cell lines originating from Ca2+ trans-
knockout mice. FASEB J 16:1398–1406, 2002
7. ASHBURNER M, BALL CA, BLAKE JA, et al: Gene ontology: Tool
porting segments are necessary to confirm the specific
for the unification of biology. The Gene Ontology Consortium. Nat
genomic regulation of particular genes.
Apart from normalizing plasma Ca2+ levels by Ca2+
8. HOENDEROP JG, DE PONT JJ, BINDELS RJ, WILLEMS PH: Hormone-
stimulated Ca2+ reabsorption in rabbit kidney cortical collect-
supplementation in 1a-OHase−/− mice, PTH levels were
ing system is cAMP-independent and involves a phorbol ester-
also normalized. Ca2+-induced effects could, therefore,
insensitive PKC isotype. Kidney Int 55:225–233, 1999
be secondary to a change in the circulating PTH lev-
9. VAN BAAL J, YU A, HARTOG A, et al: Localization and regulation by
vitamin D of calcium transport proteins in rabbit cortical collecting
els. The third comparison of Ca2+ supplementation in
system. Am J Physiol 271:F985–F993, 1996
1a-OHase+/− mice with normal 1,25(OH)2D3 and PTH
10. BINDELS RJ, RAMAKERS PL, DEMPSTER JA, et al: Role of Na+/Ca2+
levels suggests that these genes are regulated by Ca2+,
exchange in transcellular Ca2+ transport across primary culturesof rabbit kidney collecting system. Pflugers Arch 420:566–572,
irrespective of the 1,25(OH)2D3 and PTH state.
11. DARDENNE O, PRUD’HOMME J, HACKING SA, et al: Correction of
CONCLUSION
the abnormal mineral ion homeostasis with a high-calcium, high-phosphorus, high-lactose diet rescues the PDDR phenotype of
The present study demonstrated that high dietary Ca2+
mice deficient for the 25-hydroxyvitamin D-1alpha-hydroxylase
is an important regulator of Ca2+ homeostasis in a vi-
(CYP27B1). Bone 32:332–340, 2003
12. HOENDEROP JG, NILIUS B, BINDELS RJ: Molecular mechanisms of
tamin D–deficient status. Using microarray analysis, we
active Ca2+ reabsorption in the distal nephron. Ann Rev Physiol
have demonstrated that dietary Ca2+ normalizes a large
part of the renal gene expression changes resulting from
13. ANZAI N, DEVAL E, SCHAEFER L, et al: The multivalent PDZ domain-
containing protein CIPP is a partner of acid-sensing ion channel 3
the absence of 1a-OHase. We also identified novel genes
in sensory neurons. J Biol Chem 277:16655–16661, 2002
and biological pathways that are regulated in the Ca2+-
14. KURSCHNER C, MERMELSTEIN PG, HOLDEN WT, SURMEIER DJ: CIPP,
mediated rescue of 1a-OHase−/− mice. Subsequent stud-
a novel multivalent PDZ domain protein, selectively interacts withKir4.0 family members, NMDA receptor subunits, neurexins, and
ies on these potential targets will provide insight in the
neuroligins. Mol Cell Neurosci 11:161–172, 1998
molecular rescue mechanisms of dietary Ca2+ supple-
15. VAN DE GRAAF SF, HOENDEROP JG, GKIKA D, et al: Functional ex-
pression of the epithelial Ca2+ channels (TRPV5 and TRPV6)requires association of the S100A10-annexin 2 complex. Embo J22:1478–1487, 2003
ACKNOWLEDGMENTS
16. CHEVESICH J, KREUZ AJ, MONTELL C: Requirement for the PDZ
domain protein, INAD, for localization of the TRP store-operated
This work was supported by the Dutch Organization of Scientific
channel to a signaling complex. Neuron 18:95–105, 1997
Research (Zon-Mw 016.006.001, Zon-Mw 902.18.298), the Dutch Kid-
17. LI HS, MONTELL C: TRP and the PDZ protein, INAD, form the
ney Foundation (C00.1881 and NSN6013), and by the Shriners of North
core complex required for retention of the signalplex in Drosophila
America. R. St-Arnaud is a “chercheur-boursier” from the Fonds de la
photoreceptor cells. J Cell Biol 150, 2000
Recherche en Sant ´e du Qu ´ebec. The research of B.B. is supported by a
18. GOEL M, GARCIA R, ESTACION M, SCHILLING WP: Regulation of
fellowship of the Dutch Academy of Arts and Sciences. Drosophila TRPL channels by immunophilin FKBP59. J Biol Chem276:38762–38773, 2001
Reprint requests to Ren´e J.M. Bindels, 160 Cell Physiology, Nijmegen
19. SIEKIERKA JJ, HUNG SH, POE M, et al: A cytosolic binding protein
Centre for Molecular Life Sciences, University Medical Centre Nijmegen,
for the immunosuppressant FK506 has peptidyl-prolyl isomerase
P.O. Box 9101, NL-6500 HB Nijmegen, The Netherlands.
activity but is distinct from cyclophilin. Nature 341:755–757, 1989
20. HARDING MW, GALAT A, UEHLING DE, SCHREIBER SL: A recep-
tor for the immunosuppressant FK506 is a cis-trans peptidyl-prolyl
REFERENCES
isomerase. Nature 341:758–760, 1989
21. AICHER L, MEIER G, NORCROSS AJ, et al: Decrease in kid-
1. DARDENNE O, PRUD’HOMME J, ARABIAN A, et al: Targeted inac-
ney calbindin-D 28 kDa as a possible mechanism mediating cy-
tivation of the 25-hydroxyvitamin D3–1(alpha)-hydroxylase gene
closporine A- and FK-506-induced calciuria and tubular mineral-
(CYP27B1) creates an animal model of pseudovitamin D-deficiency
ization. Biochem Pharmacol 53:723–731, 1997
rickets. Endocrinology 142:3135–3141, 2001
22. ASHBY MC, TEPIKIN AV: Polarized calcium and calmodulin signaling
2. PANDA DK, MIAO D, TREMBLAY ML, et al: Targeted ablation of the
in secretory epithelia. Physiol Rev 82:701–734, 2002
25-hydroxyvitamin D 1 alpha-hydroxylase enzyme: Evidence for
23. SAIMI Y, KUNG C: Calmodulin as an ion channel subunit. Annu Rev
skeletal, reproductive, and immune dysfunction. Proc Natl Acad
24. ZUHLKE RD, PITT GS, DEISSEROTH K, et al: Calmodulin supports
3. SONG Y, KATO S, FLEET JC: Vitamin D receptor (VDR) knockout
both inactivation and facilitation of L-type calcium channels. Nature
mice reveal VDR-independent regulation of intestinal calcium ab-
sorption and ECaC2 and calbindin D9k mRNA. J Nutr 133:374–380,
25. BOULAY G: Ca2+-calmodulin regulates receptor-operated Ca2+ en-
try activity of TRPC6 in HEK-293 cells. Cell Calcium 32:201–207,
4. AMLING M, PRIEMEL M, HOLZMANN T, et al: Rescue of the skele-
tal phenotype of vitamin D receptor-ablated mice in the setting of
26. NIEMEYER BA, BERGS C, WISSENBACH U, et al: Competitive regu-
normal mineral ion homeostasis: Formal histomorphometric and
lation of CaT-like-mediated Ca2+ entry by protein kinase C and
biomechanical analyses. Endocrinology 140:4982–4987, 1999
calmodulin. Proc Natl Acad Sci USA 98:3600–3605, 2001
5. VAN CROMPHAUT S, DEWERCHIN M, HOENDEROP JG, et al: Active
27. DOLMETSCH RE, PAJVANI U, FIFE K, et al: Signaling to the nucleus by
duodenal calcium absorption in vitamin D receptor-knock out mice:
an L-type calcium channel-calmodulin complex through the MAP
functional and molecular aspects. Proc Natl Acad Sci USA 98:13324–
kinase pathway. Science 294:333–339, 2001
28. KIM E, NIETHAMMER M, ROTHSCHILD A, et al: Clustering of Shaker-
6. HOENDEROP JG, DARDENNE O, VAN ABEL M, et al: Modulation
type K+ channels by interaction with a family of membrane-
of renal Ca2+ transport protein genes by dietary Ca2+ and 1,25-
associated guanylate kinases. Nature 378:85–88, 1995
INFORMAZIONI UTILI RELATIVE AL VIAGGIO CON IL TRENO Ciascun passeggero sarà dotato di una tessera che permetterà l’addebito dei servizi a pagamento (bar, bevande, lavanderia, telefono ecc.) in un unico conto. I passeggeri potranno saldare tali servizi di volta in volta o addebitarli su un conto da saldare alla fine del viaggio in contanti o con carta di credito. I conti di importo in
FOR IMMEDIATE RELEASE Contact:Eric Eddydouble E communications212-941-7590email: [email protected] WILD BRAIN ANIMATES LUNCHABLES CAMPAIGN FOR KRAFT FOODS VIA J. WALTER THOMPSON Award-Winning Animation Studio Creates New ‘Lunchables Brigade’ Icons; Treatment of 3D Animé Characters with Flat Color Rendering A Fresh Look for Category SAN FRANCISCO, CA – March 1
 Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
containing 25% (wt/vol) formamide (Bio-Rad Labora-tories), 5× SSC [sodium chloride) (Riedel-de Ha ¨en)];
sodium citrate dihydrate (Merck, Dramstadt, Germany);
0.1% (wt/vol) SDS; 200 lg/mL of Herring Sperm DNA(Invitrogen Life Sciences); and tRNA (Roche Molecular
Diagnostics GmbH, Mannheim, Germany) that had beendenatured at 95◦C for 5 minutes and incubated at 42◦C
overnight in acclimatized hybridization chambers (Corn-
ing Incorporated Life Sciences, Acton, MA, USA). Slides
were then washed first, in 100 mL 1× SSC with 0.2%(wt/vol) SDS; second, in 0.1× SSC with 0.2% (wt/vol)
Scanning/quantification of cDNA chips
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
containing 25% (wt/vol) formamide (Bio-Rad Labora-tories), 5× SSC [sodium chloride) (Riedel-de Ha ¨en)];
sodium citrate dihydrate (Merck, Dramstadt, Germany);
0.1% (wt/vol) SDS; 200 lg/mL of Herring Sperm DNA(Invitrogen Life Sciences); and tRNA (Roche Molecular
Diagnostics GmbH, Mannheim, Germany) that had beendenatured at 95◦C for 5 minutes and incubated at 42◦C
overnight in acclimatized hybridization chambers (Corn-
ing Incorporated Life Sciences, Acton, MA, USA). Slides
were then washed first, in 100 mL 1× SSC with 0.2%(wt/vol) SDS; second, in 0.1× SSC with 0.2% (wt/vol)
Scanning/quantification of cDNA chips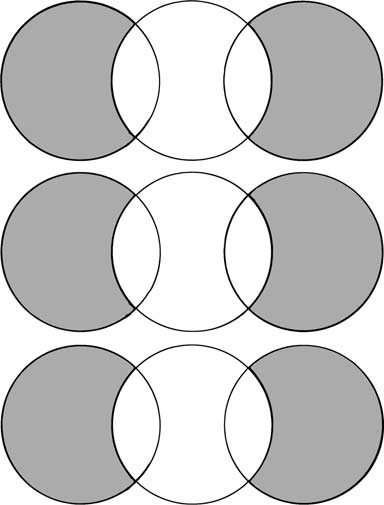
 Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Shared −/− vs. −/− Shared +/− vs. +/−
Fig. 2. Number of genes regulated when comparing the three condi-
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Shared −/− vs. −/− Shared +/− vs. +/−
Fig. 2. Number of genes regulated when comparing the three condi-
 Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Analysis using GO Consortium functional
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Analysis using GO Consortium functional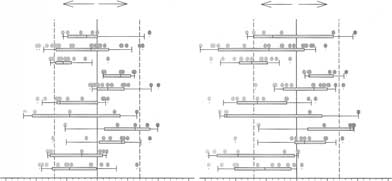 Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Fig. 5. A subset of regulated functional gene
Hoenderop et al: Gene profiling of 1a-hydroxylase knockout mice
Fig. 5. A subset of regulated functional gene