Veterinary Microbiology 95 (2003) 75–89
Interleukin 6, serum amyloid A and haptoglobin
experimentally infected with Actinobacillus
C. Hultén , E. Johansson , C. Fossum , P. Wallgren
a Department of Clinical Chemistry, P.O. Box 7038, Uppsala, Sweden
b Division of Immunology, Department of Veterinary Microbiology, BMC,
c Department of Large Animal Clinical Sciences, Faculty of Veterinary Medicine,Swedish University of Agricultural Sciences, P.O. Box 7018,
d National Veterinary Institute, SE-751 89 Uppsala, Sweden
Received 17 October 2002; received in revised form 18 March 2003; accepted 28 March 2003
Abstract
The possibility to use acute phase proteins to monitor the elimination of a bacterial infection in
pigs would facilitate an objective assessment of treatment with various antimicrobial substances. Toexamine this possibility, the acute phase response (IL-6, serum amyloid A (SAA), and haptoglobin)elicited by Actinobacillus pleuropneumoniae and its reduction on treatment with various antibioticswas studied in serum from specific pathogen free (SPF) pigs. Pigs were infected intranasally withA. pleuropneumoniae serotype 2, and either left as non-treated control pigs or treated with differ-ent antibiotics intramuscularly at onset of respiratory disease (20 h post-infection). Pigs respondedto the infection with prominent increases in activity and concentrations of IL-6, SAA, and hap-toglobin. These responses were to a certain extent overlapping and covered the time span from afew hours after infection until development of detectable levels of specific antibodies (7–10 dayspost-infection in untreated pigs). The haptoglobin response lasted until the end of the study on day17 and thereby partly coincided with the antibody response. Treatment with antimicrobials that effec-tively reduced establishment of the infection with A. pleuropneumoniae also reduced the duration ofall three acute phase responses, and reduced the concentration of serum haptoglobin. In contrast, lessefficacious treatments did not reduce these acute phase responses. Thus, acute phase reactants can beapplied to monitor therapeutic effects of antimicrobial drugs in the pig and measurements of IL-6,
∗ Corresponding author. Tel.: +46-18-67-40-00; fax: +46-18-30-91-62. E-mail address: [email protected] (P. Wallgren).
0378-1135/03/$ – see front matter 2003 Elsevier Science B.V. All rights reserved. doi:10.1016/S0378-1135(03)00136-6
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
SAA and haptoglobin could add valuable information about the stage of infection during a diseaseoutbreak. 2003 Elsevier Science B.V. All rights reserved. Keywords: Pig; Bacteria; Actinobacillus pleuropneumoniae; Antibiotics; IL-6; Serum amyloid A; Haptoglobin
1. Introduction
Diseases of the respiratory tract constitute a major health disturbance within pig pro-
duction and one of the dominating causative agents within thisdisease complex is Actinobacillus pleuropneumoniae. The bacteria may, alone or in com-bination with other microorganisms, cause considerable harm to affected pigs. The acuteform of actinobacillosis, that may be life threatening, is manifested by a sudden onsetof severe respiratory signs that coincide with increased body temperature and anorexia. However, a chronic onset of disease is more common and the infection can even proceedsubclinically (Being a Gram-negative rod, A. pleuropneumoniae containslipopolysaccharides (LPS), which are potent inducers of the acute phase response in severalspecies () but also otherbacterial compounds such as fimbriae, muramyl dipeptide and peptidoglycan can inducethe production of pro-inflammatory cytokines (Furthermore,A. pleuropneumoniae invasion of lungs causes tissue damage that may contribute to the acute phase response.
The prompt elevation of cytokine and acute phase protein concentrations in serum follow-
ing microbial invasion makes these proteins useful as inflammatory markers in veterinarymedicine In the pig, several acute phase proteins, including haptoglobin (C-reactive protein (CRP), major acute phase protein (pig-MAP)and serum amyloid A (SAA) have been proven potentially use-ful as inflammatory markers of A. pleuropneumoniae infections. In addition, productionof cytokines, such as TNF-α, IL-1 (and IL-6 (has been demonstrated in serum following challenge with A. pleu-ropneumoniae. A. pleuropneumoniae is generally susceptible to several antimicrobial agents as deter-
mined in vitro, and consequently different treatment regimens of diseased pigs have beenapplied. Substances reported to be effective include penicillin G enrofloxacin (cephalosporins and tia-mulin (Evaluation of treatment with these antimicrobialsusing recordings of clinical signs of disease, development of antibodies and pathologicallesions at autopsy have, however, indicated different efficacy (Con-sequently, a relationship between the efficacy of therapy and the magnitude of acute phaseprotein responses mounted following an infection with A. pleuropneumoniae would beexpected.
In the present study, three acute phase markers (IL-6, SAA and haptoglobin) were
determined in serum from pigs experimentally infected with A. pleuropneumoniae and
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
subsequently treated with different antibiotic substances. The magnitude and duration ofthe acute phase responses were compared and related to clinical observations in order toevaluate their usefulness in the assessment of treatment efficacy. 2. Materials and methods 2.1. Animals, experimental infection and antibiotic treatment
Specific pathogen free (SPF) Swedish Landrace × Swedish Yorkshire pigs (Serogrisen,
Ransta, Sweden; were purchased at the age of 9 weeks. On arrivalat the National Veterinary Institute (NVI) the pigs were allotted into groups, taking intoaccount litter origin, weight and sex. The pigs, which were allowed to acclimatise for 1week before start of the experiment, were fed a commercial dry feed diet (Slaktfor 290,Lantmännen, Svalöv, Sweden) and had free access to water.
The experiment (see three groups with 10 pigs each which were housed
in separate rooms. Eight days after arrival (day 0) all pigs in two of the groups wereinfected intranasally with 108 colony forming units (CFU) of A. pleuropneumoniae serotype2 (strain 700/89) grown overnight on PPLO agar at 37 ◦C in a humid atmosphere with5% CO2. After development of clinical signs, at 20 h post-infection (pi), pigs in one ofthe infected groups were given intramuscular injections with 2.5 mg enrofloxacin per kgbody weight once daily for 3 days (Baytril® 25 mg/ml, Bayer, Leverkusen, Germany), atreatment known to be effective in curing clinical A. pleuropneumoniae infection (Pigs in the other infected group were left untreated whereas pigs in the thirdgroup served as uninfected, untreated control pigs. The experiment was ended 17 days post-infection.
Fig. 1. Schematic description of the A. pleuropneumoniae infection model used for assessment of antibiotic therapyin pigs, for details see indicate: clinical examination (rhombs),measurement of rectal temperature (squares), blood sampling for acute phase reactants (triangles) and serumantibodies to A. pleuropneumoniae (circles), respectively. The recordings of respiratory symptoms, appetite andrectal temperature were carried out twice daily during the first 2 days of infection. During that period, bloodsamples were collected at 0 h; 10, 20, 28 and 44 h pi. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–892.2. Clinical recordings and sampling procedures
Clinical recordings and blood sampling were performed as outlined in
signs were scored grading severity of symptoms as 0 (no respiratory signs), 1 (moderatelyforced breathing and hoarseness), 2 (moderately forced breathing and sporadic cough orseverely forced breathing) and 3 (severely forced breathing and intermittent cough). Thefeed intake was graded on group basis from zero (refusal) to five (all consumed). Pigs wererestrained by snaring when rectal body temperatures were measured. Blood was collectedby jugular venipuncture using tubes without additive (Becton-Dickinson, Meylan Cedex,France). After centrifugation and collection of serum, samples were stored at −20 ◦C. 2.3. Necropsies and reisolation of A. pleuropneumoniae
All pigs were euthanized at the end of the experiment (on day 17) and the lungs were
inspected for signs of pneumonia and pleuritis. Samples for isolation of A. pleuropneumo-niae were collected with sterile cotton swabs from affected parts of the lung, bronchus,bronchial lymph node, pericardium and, when present, from abscesses. In lungs withoutlesions the sample was collected from the dorsal part of the diaphragmal lobe. The sampleswere cultured on blood agar plates and cross-inoculated with a single streak of a nurse strainof E. coli at 37 ◦C (Isolates of A. pleuropneumoniae were serotypedaccording to
2.4. Detection of antibodies to A. pleuropneumoniae
Serum samples were analysed for presence of antibodies to A. pleuropneumoniae using
an ELISA technique as previously described The sampleswere diluted 1:1000 and absorbance values exceeding 0.3 OD units when read at 450 nmwere considered as positive reactions. 2.5. Detection of interleukin-6, SAA and haptoglobin
IL-6 activity in serum was measured using a bioassay based on proliferation of cell line B9
as previously described (Levels of IL-6 activitywere calculated using a serially diluted murine recombinant IL-6 preparation (GenzymeDiagnostics, Cambridge, MA) as a laboratory standard. The detection limit of the assaywas 0.3 U/ml and the standard curve range was 0.03–5 U/ml.
Serum concentrations of SAA were measured using a commercially available kit (PhaseTM
Range SAA Assay, Tridelta Development Ltd., Greystones, Wicklow, Ireland). The as-say was performed as recommended by the manufacturer, except for the standard curverange that was extended to a working range comprising sample concentrations from 19.5to 1250 mg/l (sample dilution 1:500). Within this interval the intra- and interassay coeffi-cients of variation were <10%. Concentrations twice as high as the detection level wereconsidered as positive reactions.
Serum concentrations of haptoglobin were measured using a commercially available
kit (PhaseTM Range Haptoglobin Assay, Tridelta Development Ltd.). The haptoglobin
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
assay was performed according to the manufacturers instructions on an automated anal-yser (Coba’s Mira, Hoffmann-La Roche, Basel, Switzerland). The assay is based on acolorimetric reaction and the working range of the assay is 0.05–6 g/l. Intra- and interassaycoefficients of variation were <1.5 and <4%, respectively. The haptoglobin concentrationsabove 1.0 g/l were considered as positive reactions. 2.6. Archived serum samples from a previous experimental infection ofSPF-pigs with A. pleuropneumoniae
After evaluation of the initial experiment, the study was expanded by analysing archived
serum samples collected during a previous experimental infection with A. pleuropneumo-niae, using the same experimental model. The serum samples originated from six experi-mental groups with eight pigs in each. Pigs in one group were infected but not treated andone control group included non-infected, non-treated pigs. Pigs in the other four groups weretreated with different antibiotic substances (enrofloxacin, 2.5 mg/kg body weight Baytril®,Bayer, Leverkusen, Germany; ceftiofur, 3.0 mg/kg body weight, Exenel®, Pharmacia &Upjohn Animal Health, Kalamazoo, USA; penicillin G, 2.0 mg/kg body weight Penovet®,Boeringer-Ingelheim Vetmedica, Germany; tiamulin, 15.0 mg/kg body weight, Tiamutin®,Leo, Copenhagen, Denmark). All treatments were initiated at 20 h pi and all substanceswere administered once a day for 3 days. For a more detailed description see
The duration of clinical signs of disease was evaluated by calculating the median number
of days with respiratory symptoms and/or increased body temperatures (>40.0 ◦C), and theduration of the acute phase responses was evaluated as median number of sampling dayswith increased concentrations of IL-6 (>0.3 U/ml), SAA (>40 mg/l) or haptoglobin (>1 g/l). The magnitudes of the acute phase responses are given as mean values ± S.E.M. in the median values and the range of the peak responses are given in The summary measurements compared statistically using two-samplet-test (treated groups versus infected non-treated group). 3. Results 3.1. Clinical, bacteriological and pathological signs of infection
As seen in all pigs experimentally infected with A. pleuropneumoniae devel-
oped clinical signs of disease including respiratory symptoms, increased body temperature(>40.0 ◦C) and loss of appetite (graded as less than five). The duration of respiratory symp-toms and fever was shortened by the treatment with enrofloxacin (non-treated versus treated:P = 0.0001 and 0.002, respectively). Four of the 10 treated pigs and all of the infected butuntreated pigs seroconverted to A. pleuropneumoniae by day 17. On that day, non-treatedpigs had higher concentrations of serum antibodies to A. pleuropneumoniae (P < 0.0001)
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 2. Rectal temperature (circles), appetite (squares) and respiratory signs (triangles) on group level (medianvalues) in pigs infected with A. pleuropneumoniae and treated with enrofloxacin (open symbols) or left untreated(closed symbols). The pigs were infected on day 0 and the treated pigs were injected with enrofloxacin (2.5 mg/kgbody weight) once a day for 3 days starting at 20 h pi. Respiratory signs were scored from 0 (no signs) to 3 (severerespiratory signs). Appetite was scored from five (all consumed) to zero (refusal).
than the enrofloxacin-treated pigs; mean absorbance values 1.29 (range 0.59–1.73) versus0.25 (range 0.03–0.52). One of the treated pigs had lesions of pneumonia whereas nine ofthe infected non-treated pigs had both pneumonia and pleuritis at necropsy. A. pleuropneu-moniae could be reisolated from two treated pigs and from six non-treated pigs.
No clinical signs of disease were recorded in the control group. The control animals did
not seroconvert to A. pleuropneumoniae and had no lesions of pneumonia or pleuritis atnecropsy. A. pleuropneumoniae could not be isolated from the control pigs. 3.2. IL-6 activity and concentrations of serum amyloid A and haptoglobin
The mean IL-6 activity of all animals in the three experimental groups is shown in
As indicated by the large S.E.M. values, a considerable individual variation wasobserved. This variation was partly due to the short duration (<8 h) of the IL-6 responseand that individual pigs reached peak values at different times. Therefore, the statisticalcomparisons between groups were carried out using median peak values. IL-6 activity was
Fig. 3. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from the experimental pigs. The pigswere infected day 0 with A. pleuropneumoniae and treated with enrofloxacin (shaded triangles) or left untreated(closed squares). The control pigs (open circles) were not infected and left untreated. Treated pigs were injectedwith enrofloxacin (2.5 mg/kg body weight) once a day for 3 days starting at 20 h pi. The results are given as meanvalues ± S.E.M., n = 10. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
detected in serum from all infected but non-treated pigs (median peak value = 32.5 U/ml,range 8.5–50, n = 10). In all but one of these pigs, IL-6 was only detected at one occasionof sampling. The majority of the pigs (n = 6) were positive at 20 h pi, one pig 28 h pi,two pigs 44 h pi and one pig on day 4. Four of the infected and enrofloxacin-treated pigsdisplayed IL-6 in serum at one occasion; three of them at 20 h pi and one at 10 h pi (medianpeak value = 14 U/ml, range 3.0–40, n = 4). In the control group, IL-6 activity wasdemonstrated in serum collected from one pig 28 pi (24 U/ml).
The mean SAA concentrations in serum of animals in the three experimental groups
are illustrated in In the following text, however, the onset of the SAA response isgiven on an individual basis, and median peak concentrations are used to allow comparisonswith the kinetics and magnitude of the IL-6 response. SAA was detected in serum from allinfected but non-treated pig (median peak value = 884 mg/l, range 258–1250, n = 10). Ingeneral, SAA responses above 100 mg/l appeared somewhat later than the IL-6 response(20 h pi, n = 2; 28 h pi, n = 4; 44 h pi, n = 2; day 4, n = 2) and lasted longer, from 1 up to4 days. The highest SAA concentrations were recorded at 44 h pi (5/10) or at day 4 (3/10). SAA was demonstrated in serum from 7 out of 10 infected and enrofloxacin-treated pigs(median peak value = 572 mg/l, range 168–1250, n = 7). Among these animals, SAA wasfirst recorded 20 h pi (n = 2), 28 h pi (n = 1) and 44 h pi (n = 4) and in most cases theseSAA responses were below detectable levels within 1 day. SAA was not demonstrated inserum collected from pigs in the control group.
The mean haptoglobin concentrations in serum of animals in the three experimenal groups
are illustrated in responses above 1.0 g/l were detected in serum fromall infected non-treated pigs (median peak value = 4.5 g/l, range 3.2–5.8, n = 10). Theresponses were first observed at 20 h (n = 1), 28 h (n = 5), 44 h (n = 2) or 4 days (n =2) pi. In one pig the highest haptoglobin level was recorded at 44 h pi, but most individualpeak haptoglobin levels appeared late (day 4, n = 6; day 7, n = 2; and day 13, n = 1). The haptoglobin responses among these pigs were long-lasting, from 6 to 16 days. Twoof the enrofloxacin-treated pigs did not display haptoglobin levels exceeding 1.0 g/l serumand these pigs were also negative for SAA. Among the other enrofloxacin-treated pigs,haptoglobin was first demonstrated 20 h pi (n = 2), 28 h pi (n = 5) and 44 h pi (n = 1). Peak values (median peak value = 2.3 g/l, range 1.0–2.8, n = 8) were recorded at 28 hpi (n = 1) or at 44 h pi (n = 7), and among these pigs the response only lasted for 1–3days. Thus, the haptoglobin response in the non-treated pigs lasted longer (P = 0.001) andreached higher (P < 0.001) levels than in the enrofloxacin-treated pigs. In the control group,the haptoglobin concentrations remained below 1 g/l (range 0.01–0.86 g/l) throughout theexperimental period. 3.3. Clinical effects of treatment with various antibiotics during anexperimental infection with A. pleuropneumoniae
The results above indicated that both the duration and the magnitude of the acute phase
responses to A. pleuropneumoniae were influenced by an effective antibiotic treatment. Therefore, serum samples collected from pigs infected with A. pleuropneumoniae andtreated with antimicrobial substances of varying efficacy were selected for further stud-ies. The clinical effects of the different treatment regimens have previously been described
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Table 1Clinical, bacteriological and pathological findings in pigs experimentally infected with A. pleuropneumoniae(A. pp) and treated with various antimicrobial
a The results (mean values ±S.D.) are summarised from previously published data
in detail and are briefly summarized in Treatment withenrofloxacin and ceftiofur were effective whereas penicillin and tiamulin less efficientlyeliminated the bacteria, as determined by cultivation and development of antibodies to A. pleuropneumoniae. 3.4. Effects of treatment with different antibiotic substances on the acute phaseresponse to A. pleuropneumoniae
The archived serum samples had previously been analysed for IL-6 activity
and were now analysed for SAA and haptoglobin. In general, infection withA. pleuropneumoniae induced an immediate IL-6 response of short duration followed bySAA and haptoglobin responses (The IL-6 response was significantlyshortened (P = 0.02) in groups treated with enrofloxacin or ceftiofur when compared withthe infected control group.
Increased levels of SAA were observed within 24 h pi, but in all groups the peak values
were recorded at 44 h pi (Four days post-infection, SAA could still be detected inserum from some pigs in all infected groups, but a substantial SAA response was at thattime only observed in serum obtained from pigs treated with tiamulin or from untreatedpigs. In these groups, as well as in the group treated with penicillin, SAA was detectedin serum from one out of eight pigs also on day 7. The SAA response was significantlyshortened (P = 0.04) in groups treated with enrofloxacin or ceftiofur when compared withthe infected control group.
A slight increase in haptoglobin levels was detected in serum obtained 20 h pi, but the
peak values were reached later and the response lasted longer than for IL-6 and SAA(The haptoglobin response was highest in untreated and tiamulin treated pigs witha similar kinetic during the first 2 weeks post-infection. Pigs in the penicillin treated grouphad elevated levels of haptoglobin throughout the experimental period. In comparison, thehaptoglobin response was clearly reduced in pigs treated with enrofloxacin or ceftiofur fromthe second day pi. The haptoglobin response was significantly shortened in groups treatedwith enrofloxacin (P = 0.01) or ceftiofur (P = 0.05) when compared with the infectedcontrol group. Furthermore, the peak concentrations of haptoglobin obtained in the infected
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 4. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from pigs infected with A. pleuropneumo-niae and treated with various anibiotics. The pigs were infected on day 0 and treated once a day for 3 consecutivedays with enrofloxacin (open triangles), ceftiofur (open circles), penicillin (closed circles) or tiamulin (closedtriangles). One group of pigs was infected but not treated (closed squares) and one group served as non-infected,untreated pigs (open squares). The results are given as mean values ± S.E.M., n = 8.
Durationthe acute phase response (IL-6, SAA and haptoglobin) in serum from pigs experimentally infected with A. pleuropneumoniae and treated
with different antimicrobials for 3 consecutive days, beginning 20 h post-infection
a The duration (days) is given as median value with the range in parenthesis.
b The magnitude (peak value) is given as median value with the range in parenthesis. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
control group was higher than in groups treated with enrofloxacin (P < 0.001), ceftiofur(P < 0.001) or penicillin (P = 0.003). 3.5. Relationship between the acute phase responses and treatment effects
As judged from clinical recordings treatments with enrofloxacin or ceftiofur
were more efficient than treatments with penicillin or tiamulin. As summarized in these treatment effects were also reflected by the acute phase response. All infectedpigs that responded with IL-6 production later also produced SAA and haptoglobin. Sixinfected pigs did not mount a detectable IL-6 response. Three of these pigs were eithertreated with enrofloxacin or ceftiofur and did not show any SAA or haptoglobin responsesduring the first 2 weeks after infection. The other three pigs were treated with penicillin ortiamulin and developed SAA and haptoglobin responses that were similar to those of theirgroup mates.
The duration of the acute phase responses in pigs treated with penicillin or tiamulin was
not significantly different from that of infected, untreated pigs. In contrast, significantly(P < 0.05) shorter IL-6, SAA and haptoglobin responses were recorded for pigs treated withenrofloxacin or ceftiofur. The magnitude of the responses, however, showed large individualvariation so that significant differences between treatment groups were only obtained forhaptoglobin. In that case, the maximal concentration was significantly (P < 0.001) higherfor infected and untreated than for those treated with penicillin, enrofloxacin or ceftiofurwhereas no effect was recorded after treatment with tiamulin. 4. Discussion
Acute phase proteins have extensively been used to monitor treatment effects in both
infectious and non-infectious diseases in humans (reviewed by In the present study, effective medical treatmentsinitiated early during the course of infection with A. pleuropneumoniae in pigs reducedthe duration of all three acute phase responses measured, and also reduced the maximalconcentrations of haptoglobin. Thus, the present results indicate that acute phase responsescan be used to monitor the therapeutic effect of antimicrobial substances in the pig. Becausethe differences in acute phase response recorded for the various treatment groups were mostprominent for haptoglobin and least evident for IL-6, acute phase proteins that developcomparatively late and last for a certain period of time seem to best reflect the efficiency ofan antimicrobial substance.
In the first part of the study, sera from three experimental groups of pigs were analysed.
In accordance with previous results (the ex-perimental infection with A. pleuropneumoniae caused a rapid onset of IL-6 production innon-treated pigs, but the individual responses were short-lived.
The SAA responses were prominent during the first week of infection, appearing when
the IL-6 responses were declining. Detection of SAA during the first week of infectionwith A. pleuropneumoniae corresponded well with earlier results (In the present study, however, the SAA response was described quantitatively, and peak
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
values were registered on the second day post-infection. Thus, the porcine SAA responseresembled that described in cattle with acute respiratory disease (and horses affected by Rhodococcus equi and coincided with signs of clinical disease(
Infection with A. pleuropneumoniae has previously been shown to elicit production of
haptoglobin and the magnitude and duration of theresponse observed in the present study correspond to previous reports (Thus, the haptoglobin response is extended over time and appears to cover themore chronic period of the disease process, as previously shown in cattle with variouschronic inflammatory diseases (Similarly, serum haptoglobinconcentrations increase during growth in conventional fattening pigs without overt diseasebut not in SPF pigs (
The results of the first trial also revealed that treatment with enrofloxacin reduced the clini-
cal signs of disease and limited the development of serum antibodies to A. pleuropneumoniae(Interestingly, enrofloxacin treatment initiated 20 h post-infectionalso reduced the magnitude and duration of the acute phase reactants. This effect was mostevident for haptoglobin concentrations when non-treated and enrofloxacin-treated pigs werecompared.
Although a relation between the antimicrobial effect and the acute phase response was
observed, it should be noted that the bacteria do not need to be intact or replicating inorder to induce cytokine production. Indeed, fluoroquinolones reduce the serum levels ofIL-6 and TNF-␣ in LPS treated mice and tetracyclines can inhibit thesecretion of TNF-␣ by porcine Kupffer cells exposed to LPS in vitro (The underlying mechanisms are not clear but in vitro studies on human monocytes havedemonstrated that tetracyclines inhibit the LPS-induced TNF-␣ and IL-1 secretion on apost-transcriptional level (which could explain the anti-inflammatoryeffect of tetracyclines. Acknowledgements
This study was financed by grants from the Swedish Council for Forestry and Agricultural
Research. Barbro Höglund is acknowledged for taking excellent care of the animals. Wethank Karin Burvall, Berit Ek, Lisbeth Fuxler, Åsa Karlsson and Maria Persson for skilfullaboratory work. References
Aarden, L.A., De Groot, E.R., Schaap, O.L., Landsorp, P.M., 1987. Production of hybridoma growth factor by
human monocytes. Eur. J. Immunol. 17, 1411–1416.
Agersø, H., Friis, C., Nielsen, J.P., 1998. Penetration of amoxycillin to the respiratory tract tissues and secretions
in Actinobacillus pleuropneumoniae infected pigs. Res. Vet. Sci. 64, 251–257.
Akunda, J.K., Johnson, E., Ahrens, F.A., Kramer, T., 2001. Chlorotetracycline modulates acute phase response of
ex vivo perfused pig livers, and inhibits TNF-␣ secretion by isolated Kupffer cells. Comp. Immunol. Microbiol. Infect. Dis. 24, 81–89. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Altman, D.G., 1991. Comparing groups—continuos data. In: Practical Statistics for Medical Research. Chapman
Anderson, M.D., Williams, J.A., 1990. Effects of tiamuline base administered intramuscularly to pigs for treatment
of pneumonia associated with Actinobacillus (Haemophilus) pleuropneumoniae. Proc. Int. Pig. Vet. Soc. Congr. 11, 15.
Baarsch, M.J., Foss, D.L., Murtaugh, M.P., 2000. Pathophysiologic correlates of acute porcine pleuropneumonia.
Biberstein, E.L., Gunnarsson, A., Hurvell, B., 1977. Cultural and biochemical criteria for the identification of
Haemophilus spp from swine. Am. J. Vet. Res. 38, 7–11.
Boosman, R., Niewold, T.A., Mutsaers, C.W.A.A.M., Gruys, E., 1989. Serum amyloid A concentrations in cows
given endotoxin as an acute-phase stimulant. Am. J. Vet. Res. 50, 1690–1694.
Christensen, G., Sørensen, V., Mousing, J., 1999. Diseases of the respiratory system. In: Straw, B.E., D’Allaire,
S., Mengeling, W.L., Taylor, D.J. (Eds.), Diseases of Swine, eighth ed. Iowa State University Press, Ames,USA, pp. 913–940.
Fossum, C., Wattrang, E., Fuxler, L., Jensen, K.T., Wallgren, P., 1998. Evaluation of various cytokines (IL-6, IFN-␣,
IFN-␥ TNF-␣) as markers for acute bacterial infection in swine—a possible role for serum interleukin-6. Vet. Immunol. Immunopathol. 64, 161–172.
Gruys, E., Obwolo, M.J., Toussaint, M.J.M., 1994. Diagnostic significance of the major acute phase proteins in
veterinary clinical chemistry: a review. Vet. Bull. 64, 1009–1018.
Gunnarsson, A., Biberstein, E.L., Hurvell, B., 1977. Serological studies on porcine strains of Haemophilusparahemolyticus (pleuropneumoniae): agglutination reactions. Am. J. Vet. Res. 38, 1111–1114.
Hall, W.F., Eurell, T.H.E., Hansen, R.D., Herr, L.G., 1992. Serum haptoglobin concentration in swine naturally or
experimentally infected with Actinobacillus pleuropneumoniae. J.A.V.M.A. 201, 1730–1733.
Heegaard, P.M.H., Klausen, J., Nielsen, J.P., González-Ramón, N., Piñeiro, M., Lampraeve, F., Alava, M.A.,
1998. The porcine acute phase response to infection with Actinobacillus pleuropneumoniae. Haptoglobin,C-reactive protein, major acute phase protein and serum amyloid A protein are sensitive indicators of infection. Comp. Biochem. Physiol. 119B, 365–373.
Henderson, B., Wilson, M., 1996. Cytokine induction by bacteria: beyond lipopolysaccharide. Cytokine 8, 269–
Horadagoda, A., Eckersall, P.D., Alsemgeest, S.P.M., Gibbs, H.A., 1993. Purification and quantitative measurement
of bovine serum amyloid-A. Res. Vet. Sci. 55, 317–325.
Horadagoda, N.U., Knox, K.M., Gibbs, H.A., Reid, S.W., Horadagoda, A., Edwards, S.E., Eckersall, P.D., 1999.
Acute phase proteins in cattle: discrimination between acute and chronic inflammation. Vet. Rec. 144, 437–441.
Hsu, F.S., Chou, S.M., Fang, F.W.S., Lin, S.B.S., 1990. Effects of ceftiofur and a pleuromutilin derivate in the
treatment of experimentally induced Actinobacillus (Haemophilus) pleuropneumoniae pneumonia in swine. Proc. Int. Pig. Vet. Soc. Congr. 11, 14.
Huang, H., Potter, A.A., Campos, M., Leighton, F.A., Willson, P.J., Haines, D.M., Yates, W.D., 1999. Pathogenesis
of porcine Actinobacillus pleuropneumoniae. Part II. Roles of proinflammatory cytokines. Can. J. Vet. Res. 63, 69–78.
Hultén, C., Demmers, S., 2002. Serum amyloid A (SAA) as an aid in the management of infectious disease in the
foal: comparison with total leukocyte count, neutrophil count and fibrinogen concentration. Equine Vet. J. 34,693–698.
Johansson, E., Fossum, C., Fuxler, L., Wallgren, P., 2001. Effects of an experimental infection with Actinobacilluspleuropneumoniae on the interferon-␣ and interleukin-6 producing capacity of porcine peripheral bloodmononuclear cells stimulated with bacteria, virus or plasmid DNA. Vet. Microbiol. 79, 171–182.
lipopolysaccharide-induced death by fluoroquinolones. Antimicrob. Agents Chemother. 44, 3169–3173.
Madsen, M.E., Carnahan, K.G., Thwaits, R.N., 1995. Evaluation of pig lungs following an experimental challenge
with Actinobacillus pleuropneumoniae serotype 1 and 5 in pigs inoculated with either hemolysin protein and/orouter membrane proteins. F.E.M.S. 131, 329–335.
Murtaugh, M.P., Baarsch, M.J., Zhou, Y., Scamurra, R.W., Lin, G., 1996. Inflammatory cytokines in animal health
and disease. Vet. Immunol. Immunopathol. 54, 45–55.
Petersen, H.H., Ersbøll, A.K., Jensen, C.S., Nielsen, J.P., 2002. Serum-haptoglobin concentration in Danish
slaughter pigs of different health status. Prev. Vet. Med. 54, 325–335. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Rygg, M., Husby, G., Marhaug, G., 1993a. Differential expression of rabbit serum amyloid A genes in response
to various inflammatory agents. Scand. J. Immunol. 38, 417–422.
Shapira, L., Soskolone, W.A., Houri, Y., Barak, V., Halabi, A., Stabholz, A., 1996. Protection against endotoxic
shock and lipopolysaccharide-induced local inflammation by tetracycline: correlation with inhibition ofcytokine secretion. Infect. Immun. 64, 825–828.
Stephano, A., Diaz, C., Vazques-Rojas, F., Navarro-Fierro, R., 1988. Efficacy of a new antimicrobial (enrofloxacin)
against experimental infection with Haemophilus pleuropneumoniae in pigs. Proc. Int. Pig. Vet. Soc. Congr. 10, 95.
Syversen, P.V., Juul, J., Marhaug, G., Husby, G., Sletten, K., 1994. The primary structure of serum amyloid A
protein in the sheep: comparison with serum amyloid A in other species. Scand. J. Immunol. 39, 88–94.
Taylor, D.J., 1999. Actinobacillus pleuropneumoniae. In: Straw, B.E., D’Allaire, S., Mengeling, W.L., Taylor, D.J.
(Eds.), Diseases of Swine, eighth ed. Iowa State University Press, Ames, USA, pp. 343–354.
Van Leeuwen, M.A., Van Rijswijk, M.H., 1994. Acute phase proteins in the monitoring of inflammatory disorders.
Bailliere’s Clin. Rheumatol. 8, 531–552.
Van Reeth, K., van Gucht, S., Penseart, M., 2002. In vivo studies on cytokine involvment during acute viral
respiratory disease of swine: troublesome but rewarding. Vet. Immunol. Immunopathol. 87, 161–168.
Wallgren, P., Persson, M., 2000. Relationship between amounts of antibodies to Actinobacillus pleuropneumoniae
serotype 2 detected in blood serum and in fluids collected from muscles of pigs. J. Vet. Med. B. 47, 727–737.
Wallgren, P., Segall, T., Mörner, A.P., Gunnarsson, A., 1999. Experimental infections with Actinobacilluspleuropneumoniae in pigs. I. Comparison of five different parenteral antibiotic treatments. J. Vet. Med. B. 46, 249–260.
Whicher, J.T., Banks, R.E., Thompson, D., Evans, S.W., 1993. The measurement of acute phase proteins as disease
markers. In: Mackiewicz, A., Kushner, I., Baumann, H. (Eds.), Acute Phase Proteins. Molecular Biology,Biochemistry, and Clinical Applications. CRC Press, Boca Raton, pp. 633–650.
Willson, P.J., Osborne, A.D., 1985. Comparison of common antibiotic therapies for Haemophiluspleuropneumoniaein pigs. Can. Vet. J. 26, 312–316.
Woo, G.H., Chae, C.H., Kim, J.H., Lee, O.S., Park, J.M., 1996. Sequential pulmonary lesions induced by
Actinobacillus pleuropneumoniae serotype 5Apx toxins in pigs. J. Agri. Sci. Vet. 38, 649–662.
Israel and Jordan – a Future of Water Technology Cooperation Israel and Jordan have a common interest in water, facing a permanent shortage of this precious and essential resource. The Arava Institute for Environmental Studies (AIES) along with the Jordan University of Science & Technology (JUST) initiated a Water Seminar for water professionals, to explore the mutual water problems and lo
Case 1 You are an Advanced term Registrar and very competent, very confident and need little supervision. The staff think the world of you and regard you as the best Sandra works as a receptionist in the Practice. She does al the scanning of correspondence for the computer. With 12 doctors, the Practice is very busy and there is a lot of scanning to be done every day. Sandra has rece


 Veterinary Microbiology 95 (2003) 75–89
Interleukin 6, serum amyloid A and haptoglobin
experimentally infected with Actinobacillus
C. Hultén , E. Johansson , C. Fossum , P. Wallgren
a Department of Clinical Chemistry, P.O. Box 7038, Uppsala, Sweden
b Division of Immunology, Department of Veterinary Microbiology, BMC,
c Department of Large Animal Clinical Sciences, Faculty of Veterinary Medicine,
Swedish University of Agricultural Sciences, P.O. Box 7018,
d National Veterinary Institute, SE-751 89 Uppsala, Sweden
Received 17 October 2002; received in revised form 18 March 2003; accepted 28 March 2003
Abstract
Veterinary Microbiology 95 (2003) 75–89
Interleukin 6, serum amyloid A and haptoglobin
experimentally infected with Actinobacillus
C. Hultén , E. Johansson , C. Fossum , P. Wallgren
a Department of Clinical Chemistry, P.O. Box 7038, Uppsala, Sweden
b Division of Immunology, Department of Veterinary Microbiology, BMC,
c Department of Large Animal Clinical Sciences, Faculty of Veterinary Medicine,
Swedish University of Agricultural Sciences, P.O. Box 7018,
d National Veterinary Institute, SE-751 89 Uppsala, Sweden
Received 17 October 2002; received in revised form 18 March 2003; accepted 28 March 2003
Abstract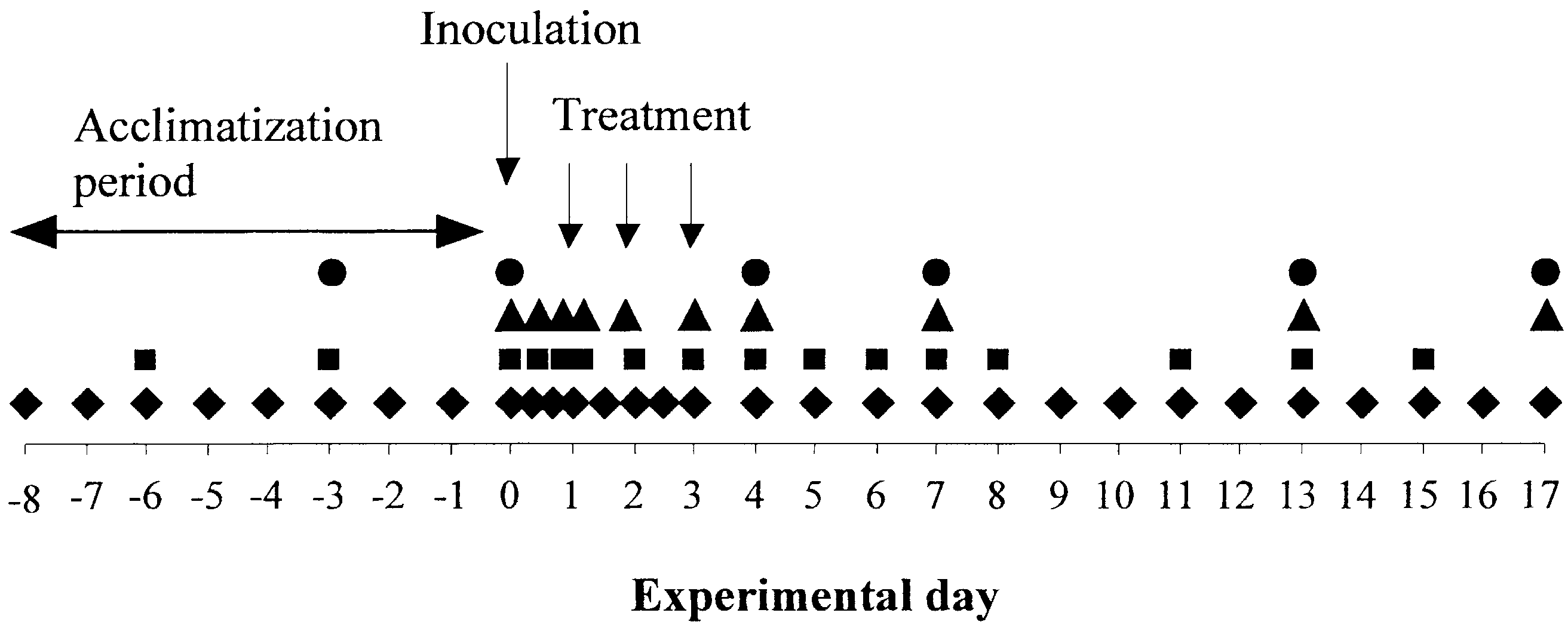 C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
subsequently treated with different antibiotic substances. The magnitude and duration ofthe acute phase responses were compared and related to clinical observations in order toevaluate their usefulness in the assessment of treatment efficacy.
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
subsequently treated with different antibiotic substances. The magnitude and duration ofthe acute phase responses were compared and related to clinical observations in order toevaluate their usefulness in the assessment of treatment efficacy. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 2. Rectal temperature (circles), appetite (squares) and respiratory signs (triangles) on group level (medianvalues) in pigs infected with A. pleuropneumoniae and treated with enrofloxacin (open symbols) or left untreated(closed symbols). The pigs were infected on day 0 and the treated pigs were injected with enrofloxacin (2.5 mg/kgbody weight) once a day for 3 days starting at 20 h pi. Respiratory signs were scored from 0 (no signs) to 3 (severerespiratory signs). Appetite was scored from five (all consumed) to zero (refusal).
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 2. Rectal temperature (circles), appetite (squares) and respiratory signs (triangles) on group level (medianvalues) in pigs infected with A. pleuropneumoniae and treated with enrofloxacin (open symbols) or left untreated(closed symbols). The pigs were infected on day 0 and the treated pigs were injected with enrofloxacin (2.5 mg/kgbody weight) once a day for 3 days starting at 20 h pi. Respiratory signs were scored from 0 (no signs) to 3 (severerespiratory signs). Appetite was scored from five (all consumed) to zero (refusal). Fig. 3. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from the experimental pigs. The pigswere infected day 0 with A. pleuropneumoniae and treated with enrofloxacin (shaded triangles) or left untreated(closed squares). The control pigs (open circles) were not infected and left untreated. Treated pigs were injectedwith enrofloxacin (2.5 mg/kg body weight) once a day for 3 days starting at 20 h pi. The results are given as meanvalues ± S.E.M., n = 10.
Fig. 3. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from the experimental pigs. The pigswere infected day 0 with A. pleuropneumoniae and treated with enrofloxacin (shaded triangles) or left untreated(closed squares). The control pigs (open circles) were not infected and left untreated. Treated pigs were injectedwith enrofloxacin (2.5 mg/kg body weight) once a day for 3 days starting at 20 h pi. The results are given as meanvalues ± S.E.M., n = 10. C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 4. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from pigs infected with A. pleuropneumo-niae and treated with various anibiotics. The pigs were infected on day 0 and treated once a day for 3 consecutivedays with enrofloxacin (open triangles), ceftiofur (open circles), penicillin (closed circles) or tiamulin (closedtriangles). One group of pigs was infected but not treated (closed squares) and one group served as non-infected,untreated pigs (open squares). The results are given as mean values ± S.E.M., n = 8.
C. Hult´en et al. / Veterinary Microbiology 95 (2003) 75–89
Fig. 4. Levels of IL-6 (a), SAA (b) and haptoglobin (c) in serum obtained from pigs infected with A. pleuropneumo-niae and treated with various anibiotics. The pigs were infected on day 0 and treated once a day for 3 consecutivedays with enrofloxacin (open triangles), ceftiofur (open circles), penicillin (closed circles) or tiamulin (closedtriangles). One group of pigs was infected but not treated (closed squares) and one group served as non-infected,untreated pigs (open squares). The results are given as mean values ± S.E.M., n = 8.