The role and source of 5P-deoxyadenosyl radical in a carbon skeleton
rearrangement catalyzed by a plant enzyme
Sandrine Ollagnier, Eric Kervio, Jaènos Reètey*
Lehrstuhl fuër Biochemie, Institut fuër Organische Chemie, Universitaët Karlsruhe, Kaiserstr. 12, D-76128 Karlsruhe, Germany
Received 15 August 1998; received in revised form 24 September 1998
Abstract The last step in the biosynthesis of tropane alkaloids
the major source of 5P-deoxyadenosyl radical [13]. More re-
is the carbon skeleton rearrangement of littorine to hyoscyamine.
cently, S-adenosylmethionine (SAM) was also identi¢ed to
The reaction is catalyzed by a cell-free extract prepared from
play a similar role [14]. Therefore, SAM has been called the
cultured hairy roots of Datura stramonium. Adenosylmethionine
stimulated the rearrangement 10^20-fold and showed saturation
Here we report on the involvement of SAM in the rear-
kinetics with an apparent Km of 25 WM. It is proposed that
rangement of littorine to hyoscyamine.
S-adenosylmethionine is the source of a 5P-deoxyadenosyl radical
which initiates the rearrangement in a similar manner as it does
in analogous rearrangements catalyzed by coenzyme B
dependent enzymes. Possible roles of S-adenosylmethionine as
a radical source in higher plants are discussed.
z 1998 Federation of European Biochemical Societies.
They were obtained from commercial sources and were the highest
purity normally available. SAM and [2,8,5P-QH]ATP (65% QH in posi-
tion 5P) were obtained from Amersham, Aylesbury, UK, dithiothreitol
Key words: Tropane alkaloid; Carbon skeleton
(DTT) from Promega, ethylene diamine tetraacetic acid (EDTA) from
rearrangement; S-Adenosylmethionine as 5P-deoxyadenosyl
Serva, B5 medium from Bioproduct (Boehringer Ingelheim).
2.2. Preparation of the phenyl-lactoyl tropine (littorine)
According to a modi¢ed procedure [16] L-(3)-phenyllactic acid
(Fluka) (116 mg, 0.7 mmol) dried over phosphorus pentoxide was
added to tropine (Acros) (0.5 mmol) previously dried over sodium
hydroxide pellets. The mixture was heated to 145³C on an oil bath
and dry hydrogen chloride was bubbled through the solution for 3.5 h.
The last and most intriguing step in the biosynthesis of the
(Note that esteri¢cation attempts with para-toluene sulfonic acid or
tropane alkaloids hyoscyamine and scopolamine is the carbon
under montmorrillonite KSF as alternative procedures gave no satis-
skeleton rearrangement depicted in Scheme 1 (for a review see
factory results.) The cooled mixture was dissolved in a 10 ml HPSOR
solution (50 mM), ¢ltered and made alkaline with a 10% aqueous
[1]). Due to its similarity to the rearrangement of methylma-
ammonium hydroxide solution, then extracted into chloroform
lonyl CoA to succinyl-CoA it was originally speculated that
(6U10 ml). The solvent was removed under reduced pressure and
the residue puri¢ed by partition chromatography on Celite (Merck)
IP is involved in the reaction [2^4]. Since, in spite
of some claims [5^7], no trace of vitamin B
containing 0.5 M phosphate bu¡er (pH 6.8) eluted with chloroform.
GC-MS analysis of the combined fractions (97 mg, yield: 48%) in-
found in plants this idea was abandoned [8,9]. More recently
dicated almost pure (98%) littorine.
it has been surmised that cytochrome P450 is the agent ab-
Retention time (1,4-dioxane) on a Optima 1-02 capillary column
stracting the benzylic hydrogen atom and redonating a hy-
(25 mU0.2 mm), ramp 100^300³C at 20³C/min): 12.2 min. MS m/z
droxyl group after the rearrangement [10,11]. Accordingly,
(rel. int.): 361(4), 193(3), 140(4), 125(12), 124(100), 123(1), 96(9),
clotrimazole, a known P-450 inhibitor, has been said to inhibit
95(5), 94(12), 91(8), 83(15), 73(19) [17].
IQC NMR and IH NMR spectra were recorded on a Bruker DRX
the formation of hyoscyamine [1], and the 3-He atom ab-
500 spectrometer. Deuteriochloroform was used as the solvent with
stracted from the benzylic position was lost during the bio-
TMS as an internal reference for IH. The abbreviations used are as
synthetic experiment [12]. When the OH group of littorine was
follows: N, shift; s, singlet; d, doublet; t, triplet; m, multiplet; b,
labelled with the isotope IVO, 25^29% of the label was lost in
the product [11]. These results were interpreted as the involve-
Q) N : 1.65 (1H, HT or HU, d, JQrr = 15.8 Hz) ; 1.85
(1H, HU or HT, d, JQrr = 15.8 Hz); 1.94^2.20 (2H, HT or HU, m); 2.00
ment of two enzymes, a mutase and a dehydrogenase, in the
(N-CHQ, s); 2.54^2.72 (4H, HR and HP, m); 2.78 (1H, HS or HI, t,
conversion of littorine into hyoscyamine, the geminal diol
JQrr = 15.8 Hz); 2.80 (1H, HI or HS, t, JQrr = 15.8 Hz); 2.98 (2H,
being an intermediate in the reaction. The dehydratation
HQH, d, JQrr = 6.5 Hz); 3.57 (b, OH); 4.37 (1H, HPH, t, JQrr = 6.5 Hz);
thereof to the aldehyde could be partially stereospeci¢c ex-
4.99 (1H, HQ, t, JQrr = 4.5 Hz); 7.11^7.22 (5Hrom, m) [18]. IQC NMR
(CDClQ) N: 25.1, 25.3 (CU and CT); 35.1, 35.2 (CR and CP); 37.3 (CQH);
plaining the less than 50% loss of IVO in the overall reaction.
40.7 (CHQ-N); 60.0 (CI and CS); 71.6 (CQ); 74.3 (CPH); 126.9 (CUH);
Only a few agents are known in biochemistry that are able to
128.5, 129.3 (CSHYTHYVHYWH); 136.4 (CRH); 173.5 (CIH).
abstract hydrogen atoms from non-activated positions. Beside
cytochrome P450, nature uses the 5P-deoxyadenosyl radical
SAM synthetase was puri¢ed by the procedure of Markham et al.
for this purpose. In animals and bacteria coenzyme BIP is
[19], with some minor modi¢cations: growth in the presence of oxy-
tetracycline (30 Wg/ml) and disruption of the cells by sonication (Ban-
delin Nonopuls HD 2200). The speci¢c activity of the enzyme was
*Corresponding author. Fax: (49) (721) 608-4823.
E-mail: [email protected]
2.4. Preparation of the S-[2,8,5P-QH]adenosylmethionine
S-[2,8,5P-QH]Adenosylmethionine was enzymatically synthesized
0014-5793/98/$19.00 ß 1998 Federation of European Biochemical Societies. All rights reserved.
PII: S 0 0 1 4 - 5 7 9 3 ( 9 8 ) 0 1 2 5 8 - 7
S. Ollagnier et al./FEBS Letters 437 (1998) 309^312
from [2,8,5P-QH]ATP (speci¢c activity: 41 mCi/mmol) and unlabelled
mercially prepared reagent (bicinchoninic acid) and bovine serum al-
methionine, using a SAM synthetase preparation.
The reaction mixture contained 200 Wl of 0.1 M diluted solution of
[2,8,5P-QH]ATP (speci¢c activity: 424 700 cpm/nmol) in 3 ml Tris-HCl
2.9. Extraction and analysis of alkaloids
bu¡er (pH 7.4), adjusted to pH 7,6 by addition of a 2 M KOH
Harvested roots were freeze-dried, then powdered and soaked over-
solution, 1.5 eq. MgClP, 5 eq. KCl, 1.2 eq. L-methionine added as
night in an EtOH-28% NHROH (19:1) mixture. This macerated ma-
solids, 100 Wl reduced glutathione (25 mg/ml) and 11 mg of SAM
terial was centrifuged for 5 min at 1300Ug. Extraction with the basic
synthetase were incubated for 45 min at 35³C. The reaction was ter-
alcohol was repeated twice and the combined alcohol fractions were
minated by addition of 40 Wl of trichloroacetic acid (6 M). The result-
evaporated to dryness at 35³C. The dry residue was dissolved in 2 ml
ing suspension was centrifuged at 4³C and the extracts pooled and
of 0.1 N HCl and the acidic aqueous solution was ¢ltered through
stored at 4³C. After lyophilization, the residue was dissolved in
¢lter No. 2. One milliliter of the ¢ltrate was made alkaline with 2 ml
doubly distilled water and applied to a carboxymethyl sepharose
of 1 M NaPCOQ-NaHCOQ bu¡er (pH 10) and 1 ml of this alkaline
high £ow column (0.8U20 cm FPLC column, Pharmacia Biotech,
solution was loaded onto a Extrelut-1 column. After 10 min, 6 ml of
Freiburg, Germany) previously equilibrated with water. The column
CHClQ was passed through the column and the chloroform fractions
was washed with 2 volumes of water and then S-[2,8,5P-
were evaporated to dryness at 35³C. The dry residue was dissolved in
QH]adenosylmethionine was eluted with 1 M NaCl gradient (4 ml/
a mixture of 1,4 dioxane/N,O-bis-(trimethylsilyl)-acetamide (v/v 4:1).
min). Fractions containing S-[2,8,5P-QH]adenosylmethionine (speci¢c
Alkaloids were measured with a gas chromatograph coupled with a
activity: 123 000 cpm/nmol) were mixed, lyophilized and stored at
mass spectrometer model Hewlett Packard 5890 Serie II using a capil-
lary column Optima 1-02 Wm (25 mU0.2 mm). The column temper-
ature was 350³C, the carrier gas was HP at a £ow rate of 20 ml/min.
Roots from Datura stramonium D15/5 were a gift from Dr. N.J.
The two alkaloids identi¢ed for this study were littorine and hyos-
Walton (Institute of Food Research, Norwich, UK). Roots cultures
were initiated by wounding surface sterilized explants of leaves on the
Littorine RT (retention time): 12.2 min. MS m/z (rel. int.): 361(4),
midrib with an overnight suspension of Agrobacterium rhizogenes.
193(3), 140(4), 125(12), 124(100), 123(1), 96(9), 95(5), 94(12), 91(8),
Tissue bearing emergent roots were excised and placed into 8 ml of
Gamborg's B5 medium supplemented by 500 Wg/ml ampicillin. Roots
Hyoscyamine RT: 12.1 min. MS m/z (rel. int.): 361(5), 193(0.5),
for enzyme extraction and puri¢cation were grown at 25³C, under
140(7), 125(10), 124(100), 123(6), 104(6), 96(10), 95(6), 94(13),
stirring at 90 rpm. Rapidly growing roots were subcultured every
2 weeks into 50 ml of the same medium in a 250 ml Erlenmeyer £ask.
After eight subcultures it should be possible to omit ampicillin from
the medium. After this time, roots were harvested by ¢ltration in
An Incapharm 100 RP-18 TS (250U4.6 mm) column was equili-
vacuo, washed twice with water and frozen immediately at 370³C.
brated at room temperature with 0.1% tri£uoroacetic acid in water
solution. 5 min after sample injection (20 Wl in same bu¡er), a gra-
dient of 0^60% solvent B (0.1% tri£uoroacetic acid in acetonitrile) in
All procedures were done at 4³C. Roots (6 g) were crushed in a
40 min was developed by a Hewlett-Packard 1050 T system at a £ow
mortar with sea sand, mixed with 15 ml of extraction bu¡er (100 mM
rate of 1.2 ml/min. The eluted substances were monitored at 215 and
KPHPOR pH 8, 3 mM DTT, 5 mM NaP EDTA and 250 mM sucrose)
254 nm. Labelled products were analyzed by liquid scintillation count-
and then ground during 30 min until a homogeneous suspension was
ing (Tri-carb 2100TR, Packard, a Canberra company, Meriben, USA)
obtained. The homogenate was clari¢ed by passage through Mira-
after adding 15 ml of scintillation liquid (Lumasafe plus^Lumac LSC)
cloth (Calbiochem, Novabiochem, Frankfurt a/M, Germany) and cen-
trifugation (30 000Ug, 4³C, 30 min). Low molecular weight com-
pounds were removed using a PD-10 desalting column (Pharmacia
2.11. Determination of the isotope exchange
Biotech AB, Uppsala, Sweden) equilibrated in a one tenth strength
Tritium exchange with water was measured by liquid scintilla-
extraction bu¡er. The proteins were eluted with 3.5 ml of the same
tion counting (Tri-carb 2100 TR) after loading 10 ml of scintillation
bu¡er. The yellow solution obtained was concentrated to 2^4 mg/ml
liquid to 10 Wl of water resulting from bulb-to-bulb distillations. The
with centricon 5000 Da (Pharmacia Biotech AB, Uppsala, Sweden).
QH]adenosylmethionine solution (speci¢c activity: 123 000 cpm/nmol)
and the resulting distilled water and between aqueous phase (0.1%
The activity of the enzyme was determined by measuring the con-
tri£uoroacetic acid in doubly distilled water) before HPLC analysis
version of littorine into hyoscyamine by the GC-MS method. The
standard reaction mixture, in a volume of 200 Wl, contained 200 WM
SAM (unlabelled or labelled as prepared above), 100 Wl of the protein
solution (200^400 Wg), 2 mM of littorine, adjusted to 200 Wl with the
extraction bu¡er. After 40 min incubation at 33³C, the reaction was
terminated by addition of 100 Wl 30% NHROH. Control experiments
3.1. Dependence of the rearrangement littorine/hyoscyamine on
lacking substrate were routinely included. Reaction mixtures were
loaded on an Extrelut-1 column (Merck Art 1.13076, Darmstadt,
Germany). After 15 min the alkaloids were eluted with 13 ml of
The experiments were carried out as described in Section
CHClQ. The chloroform fractions were evaporated to dryness at
2.7 and are listed in Table 1. The silylated products were
35³C, dissolved in 200 Wl of a fresh solution of 1,4 dioxane/N,O-bis-
detected by GLC coupled with MS. Omission of SAM from
(trimethylsilyl)-acetamide (4:1 v/v), analyzed by GC-MS (experiments
the assay mixture resulted in only 3.5% hyoscyamine while
96.5% remained littorine. Addition of SAM (200 WM) in-
creased the conversion leading to 61% hyoscyamine. When
Protein was assayed by the method of Smith et al. [20] using com-
the hairy root extract was kept frozen at 320³C for a week
Dependence of the rearrangement of tropane alkaloids on SAMHairy roots
S. Ollagnier et al./FEBS Letters 437 (1998) 309^312
The 10^20-fold enhancement of the carbon skeleton rear-
rangement, littorine to hyoscyamine (Scheme 1) by SAM in
cell-free extracts of a root culture of Datura stramonium sug-
gests a new role for this cofactor in plant biochemistry. The
well-known function of SAM as methyl donor is operative in
both prokaryotic and eukaryotic organisms and requires a
heterolytic cleavage of a carbon sulfur bond. More recently,
Scheme 1. Carbon skeleton rearrangement of tropane alkaloids.
Frey and coworkers discovered and documented [14,15] a case
in which SAM is the source of a 5P-deoxyadenosyl radical, the
homolytic cleavage of the carbon sulfur bond being promoted
the conversion under the same conditions without and with
reductively by an FeRSR cluster-containing enzyme.
SAM decreased to 2.3 and 28% hyoscyamine, respectively.
We surmise a similar role for SAM in the rearrangement of
After 4 weeks of freezing little if any mutase activity of the
littorine to hyoscyamine. Indeed, addition of SAM to the
assay mixture not only increased the conversion of littorine
to hyoscyamine, but showed saturation kinetics with a Km of
3.2. Determination of the Michaelis constant for SAM
As the enzyme source we used a hairy root extract, thawed
The low activity of the hairy root extracts without addition
after 1 week in the deep-freeze. The kinetic experiments were
of SAM may be due to trace amounts of endogenous SAM
carried out under conditions of the enzyme assay (see Section
that remained in the extract after the PD-10 desalting column.
2.7), except that the concentration of SAM was varied be-
Indeed, in previous studies no SAM was added to the cell-free
tween 50 and 500 WM. The Km and Vmx values were deter-
system, but much more extract and longer incubation times
mined using Lineweaver-Burk plot (Fig. 1).
were used. Moreover, most of the incorporation experiments
The following values were found: Km = 25 þ 5 WM, Vmx =
were carried out with growing root cultures and the incuba-
tion times were 10^28 days. In these cases endogenous SAM
must have been provided by the in vivo system.
3.3. Conducting the rearrangement in the presence of
Preliminary experiments using [2,8,5P-QH]SAM showed no
incorporation of tritium into littorine or hyoscyamine but
When the enzymic rearrangement of littorine to hyoscy-
some loss of the radioactivity to water. This is in agreement
amine was carried out in the presence of [2,8,5P-QH]SAM
with the observed loss of the benzylic pro-R H-atom of littor-
(see Section 2.4 for preparation), no incorporation of tritium
ine during the rearrangement [12]. Since all work has been
into the alkaloids was observed. However, about 2% of the
done with root cultures or crude cell-free extracts it is possible
tritium (calculated for the 5P position) was washed out into
that the migrated H-atom is washed out by redox enzymes.
the medium. Only a trace of tritium was found in the solvent
This could also explain the 25^29% loss of IVO of littorine
in a control experiment without hairy root extract. HPLC
during the rearrangement [11] since the intermediate aldehyde
showed no appreciable amounts of 5P-deoxyadenosine after
could be in equilibrium with its hydrate.
the reaction. If the latter was an intermediate, then only in
The steric course of the migration at the relevant centers is
enzyme-bound catalytic amounts. If the putative 5P-deoxyade-
also of importance. After contradictory results [2,3,21] it has
nosyl radical abstracted the benzylic pro-R H-atom, then this
been established that the substitution occurs with inversion at
process must have been irreversible. Possible reasons for
both migration centers [12]. The same steric course has been
the loss of tritium from the 5P position of SAM are given
determined for the lysine 2,3-aminomutase reaction [22]. In
the latter, however, the migrating hydrogen atom is not lost
Fig. 1. Lineweaver-Burk determination of Km and Vmx. X = % GC-MS hyoscyamine.
S. Ollagnier et al./FEBS Letters 437 (1998) 309^312
to the medium. This discrepancy can only be clari¢ed when
[8] Freer, I., Pedrochi-Fantoni, G., Picken, D.J. and Overton, K.H.
pure littorine mutase is available. Another plant enzyme that
(1981) J. Chem. Soc. Chem. Commun. 80^82.
catalyzes the rearrangement of K-phenylalanine to L-phenyl-
[9] Leete, E. (1990) Planta Med. 56, 339^352.
[10] Robins, R.J., Chesters, N.C.J.E., O'Hagan, D., Parr, A.J., Wal-
alanine is the phenylalanine aminomutase. This reaction is
ton, N.J. and Woolley, J.G. (1995) J. Chem. Soc. Perkin Trans.
part of taxol biosynthesis. Recently it has been established
that the substitution occurs with retention of con¢guration
[11] Wong, C.W., Hamilton, J.T.G., O'Hagan, D. and Robins, R.J.
at both migration centers [23]. This fact does not rule out
(1998) J. Chem. Soc. Chem. Commun. 1045^1046.
[12] Chesters, N.C.H.E., Walker, K., O'Hagan, D. and Floss, H.G.
the possibility that the same cofactor, namely SAM, is oper-
(1996) J. Am. Chem. Soc. 118, 925^926.
ative also in the phenylalanine aminomutase reaction. Prece-
[13] Frey, P.A., Essenberg, M.K. and Abeles, R.H. (1969) J. Biol.
dence for opposite steric courses is found in the coenzyme
[14] Chang, C.H., Ballinger, M.D., Reed, G.H. and Frey, P.A. (1996)
IP-dependent carbon skeleton rearrangements. Methylma-
lonyl-CoA mutase operates with retention [24,25], methylene
Biochemistry 35, 11081^11084 and previous papers cited therein.
[15] Frey, P.A., Reed, G.H., Moss, M.L., Petrovich, R.M., Ballinger,
glutarate mutase [26] and glutamate mutases [27] with inver-
M.D., Lieder, K.W., Wu, W., Chang, C.H., Bandarian, V., Ru-
sion of con¢guration. Even the same enzyme, the coenzyme
zicka, F.J., Lo Brutto, R. and Beinert, H. (1998) in: Vitamin BIP
IP-dependent ethanolamine ammonia-lyase, operates with
IP-Proteins (Kraëutler, B., Arigoni, D. and Golding, B.T.,
opposite steric courses, inversion and retention with (2R)-
[16] Robins, R.J., Bachman, P. and Woolley, J.G. (1994) J. Chem.
and (2S)-propanolamines, respectively [28]. We thus propose
that SAM may be the cofactor in the BIP-like rearrangements
[17] Hashimoto, T., Yukimune, Y. and Yamada, Y. (1986) J. Plant
occurring in plants and the reaction is initiated by the 5P-
deoxyadenosyl radical derived from SAM.
[18] Cannon, J.R., Joshi, K.R., Meehan, G.V. and Williams, J.R.
That plants use SAM as the source of 5P-deoxyadenosyl
[19] Markham, G.D., Hafner, E.W., Tabor, C.W. and Tabor, H.
radical has been established for the last step of biotin biosyn-
(1983) Methods Enzymol. 94, 219^222.
thesis in Arabidopsis thaliana [29]. A similar mechanism oper-
[20] Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gart-
ates in Bacillus sphaericus and Escherichia coli [30,31]. We
ner, F.H., Prorenzano, M.D., Fujimoto, E.K., Goeke, N.M.,
thus believe that while bacteria are able to use both coenzyme
Olson, B.J. and Klenk, D.C. (1985) Anal. Biochem. 150, 76^85.
[21] Platt, R.V., Opie, C.T. and Haslam, E. (1984) Phytochemistry 23,
BIP and SAM as sources of 5P-deoxyadenosyl radical, animals
and humans inherited from them the former and higher plants
[22] Aberhart, D.J., Gould, S.J., Lin, H.J., Thiruvengadam, T.K. and
Weiller, B.H. (1983) J. Am. Chem. Soc. 105, 5461^5470.
[23] Walker, K.D. and Floss, H.G. (1998) J. Am. Chem. Soc. 120,
Acknowledgements: The work was supported by the European Union
and the Fonds der Chemischen Industrie. We thank Dr. Nick Walton,
[24] Sprecher, M., Clark, M.J. and Sprinson, D.B. (1964) Biochem.
Institute of Food Research, Norwich, UK, and Professor Meinhart
Zenk, University of Munich, Germany, for help with establishing the
[25] Reètey, J. and Zagalak, B. (1973) Angew. Chem. Int. Ed. Engl. 12,
[26] Hartrampf, G. and Buckel, W. (1986) Eur. J. Biochem. 156, 301^
[27] Sprecher, M., Switzer, R.L. and Sprinson, D.B. (1966) J. Biol.
[1] O'Hagan, D. and Robins, R.J. (1998) Chem. Soc. Rev. 27, 207^
[28] Diziol, P., Haas, H., Reètey, J., Graves, S.W. and Babior, B.M.
(1980) Eur. J. Biochem. 106, 211^224.
[2] Leete, E. (1984) J. Am. Chem. Soc. 106, 7271^7272.
[29] Baldet, P., Alban, C. and Douce, R. (1997) FEBS Lett. 419, 206^
[3] Leete, E. (1987) Can. J. Chem. 65, 226^228.
[4] Ansarin, M. and Woolley, J.G. (1995) J. Chem. Soc. Perkin
[30] Guianvarc'h, D., Florentin, D., Tse Sum Bui, B., Nuzi, F. and
Marquet, A. (1997) Biochem. Biophys. Res. Commun. 236, 402^
[5] Fries, L. (1962) Physiol. Plant. 15, 566.
[6] Poston, J.M. (1977) Science 195, 301^302.
[31] Shaw, N.M., Birch, O.M., Tinschert, A., Venetz, V., Dietrich, R.
[7] Poston, J.M. (1978) Phytochemistry 17, 401^402.
and Savoy, L.-A. (1998) Biochem. J. 330, 1079^1085.
Overzicht Aandoeningen en Stofnamen in het kader van de Compensatieregeling Eigen Risico 2008 Verzekerden die lijden aan (een van de) onderstaande aandoeningen ontvangen een financiële compensatie voor het verplicht eigen risico. Dit geldt alleen wanneer zij minimaal 180 dagen per jaar een medicijn voorgeschreven krijgen voor één specifieke aandoening met daarin (een van de) stoffen d
 The role and source of 5P-deoxyadenosyl radical in a carbon skeleton
rearrangement catalyzed by a plant enzyme
Sandrine Ollagnier, Eric Kervio, Jaènos Reètey*
Lehrstuhl fuër Biochemie, Institut fuër Organische Chemie, Universitaët Karlsruhe, Kaiserstr. 12, D-76128 Karlsruhe, Germany
Received 15 August 1998; received in revised form 24 September 1998
Abstract The last step in the biosynthesis of tropane alkaloids
the major source of 5P-deoxyadenosyl radical [13]. More re-
is the carbon skeleton rearrangement of littorine to hyoscyamine.
The role and source of 5P-deoxyadenosyl radical in a carbon skeleton
rearrangement catalyzed by a plant enzyme
Sandrine Ollagnier, Eric Kervio, Jaènos Reètey*
Lehrstuhl fuër Biochemie, Institut fuër Organische Chemie, Universitaët Karlsruhe, Kaiserstr. 12, D-76128 Karlsruhe, Germany
Received 15 August 1998; received in revised form 24 September 1998
Abstract The last step in the biosynthesis of tropane alkaloids
the major source of 5P-deoxyadenosyl radical [13]. More re-
is the carbon skeleton rearrangement of littorine to hyoscyamine.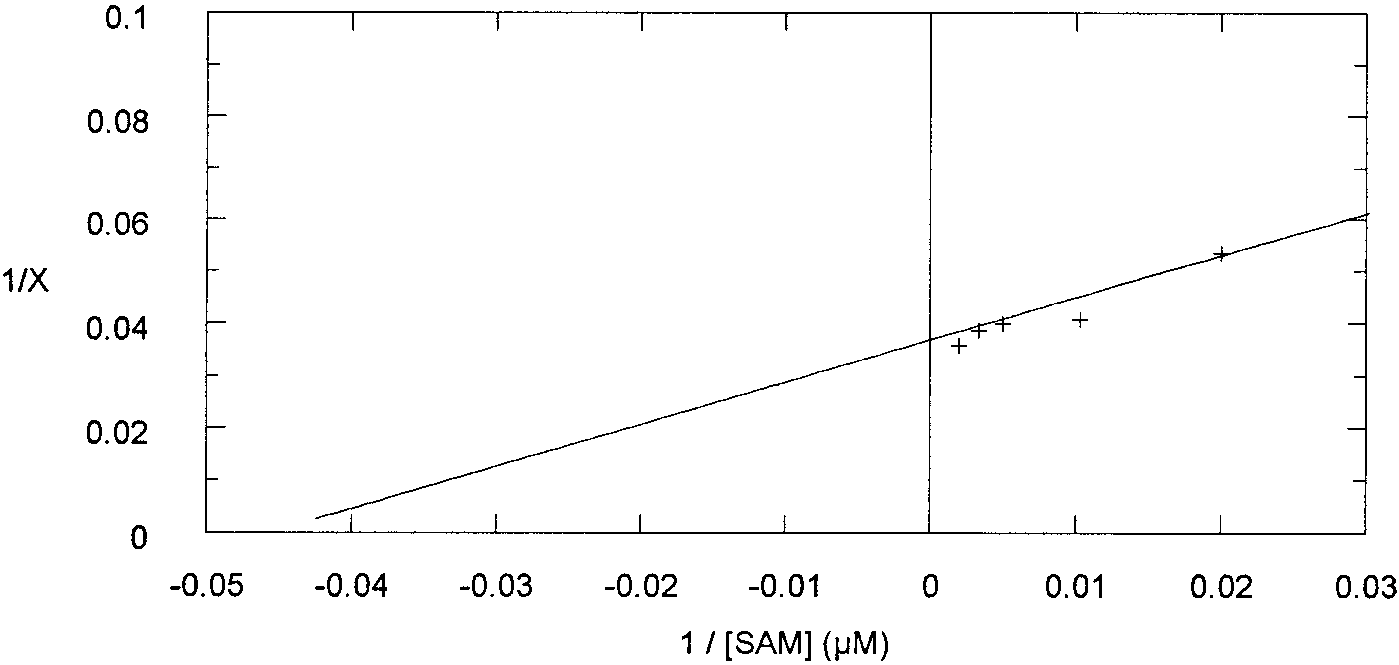
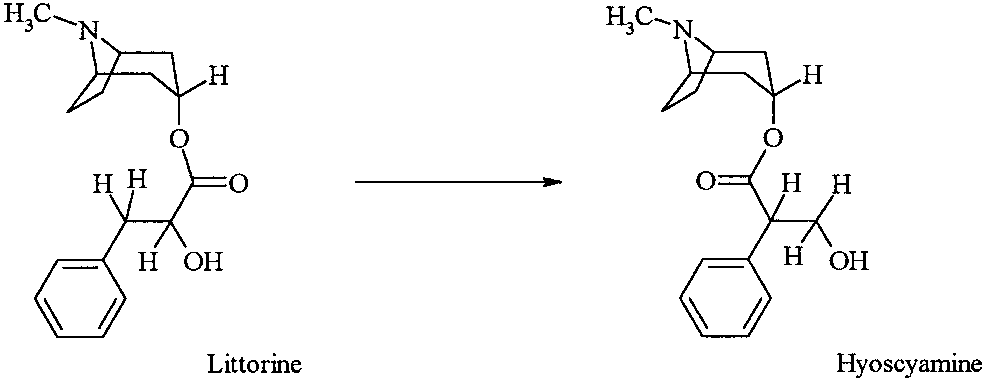 S. Ollagnier et al./FEBS Letters 437 (1998) 309^312
The 10^20-fold enhancement of the carbon skeleton rear-
rangement, littorine to hyoscyamine (Scheme 1) by SAM in
cell-free extracts of a root culture of Datura stramonium sug-
gests a new role for this cofactor in plant biochemistry. The
well-known function of SAM as methyl donor is operative in
both prokaryotic and eukaryotic organisms and requires a
heterolytic cleavage of a carbon sulfur bond. More recently,
Scheme 1. Carbon skeleton rearrangement of tropane alkaloids.
S. Ollagnier et al./FEBS Letters 437 (1998) 309^312
The 10^20-fold enhancement of the carbon skeleton rear-
rangement, littorine to hyoscyamine (Scheme 1) by SAM in
cell-free extracts of a root culture of Datura stramonium sug-
gests a new role for this cofactor in plant biochemistry. The
well-known function of SAM as methyl donor is operative in
both prokaryotic and eukaryotic organisms and requires a
heterolytic cleavage of a carbon sulfur bond. More recently,
Scheme 1. Carbon skeleton rearrangement of tropane alkaloids.